- 移动端


武汉华美生物工程有限公司CUSABIO®品牌商
15 年
手机商铺
- NaN
- 0
- 0
- 2
- 2

推荐产品

公司新闻/正文
CTRAM:I 类限制性 T 细胞相关分子,免疫球蛋白超家族(Ig-SF)新成员!
1806 人阅读发布时间:2023-12-12 17:29
CRTAM 是一种新鉴定的免疫球蛋白超家族(Ig-SF)新成员。Ig-SF 蛋白家族包含多个免疫调节和细胞粘附分子,这些蛋白在免疫系统的形成、应答和细胞交互中扮演重要角色。CRTAM 作为一种细胞表面跨膜蛋白,目前针对这一靶点的研究报道较少。相关研究表明 CRTAM(CD355)与多种疾病关联,包括炎症、哮喘、恶性间皮瘤和某些肿瘤疾病。CRTAM 促进促炎细胞因子的产生,可能参与自身免疫性疾病的免疫病理学。此外,CRTAM 在免疫过程中对 NK 细胞杀伤肿瘤细胞起到关键作用,呈现出潜在的肿瘤靶向策略。今天,我们一起了解下这个 Ig-SF 新成员 CRTAM!
1. 什么是 CTRAM?
1.1 CTRAM 的结构
CRTAM(Class I-restricted T cell-associated molecule,又称 CD355)是 I 类限制性 T 细胞相关分子。CRTAM 最初被发现在活化的 CD8+T 细胞和 NKT 细胞中表达较高,这些细胞受 I 类主要组织相容性复合物(MHC)的限制,CRTAM 因此得名。人的 CRTAM 全长有 393 个氨基酸,它的胞外区有两个免疫球蛋白结构域,第一个是 IgV,第二是 IgC。CRTAM 胞内有类-I PDZ(ESIV)结合基序,能连接到支架蛋白的 PDZ 区域。CRTAM 的两个 Ig 样结构域使其被归类免疫球蛋白超家族(Ig-SF)的一个新成员。另有报道 CRTAM 还与 Necl 蛋白家族有关,其氨基酸序列与 Necl 蛋白相似度约为 20%。因此,研究者又将 CRTAM 归类为 Nectin 样家族的成员 [1-3]。
1.2 CTRAM 的表达和功能
CRTAM 作为一种新鉴定的在 T 细胞激活标记物,其在体外激活的 iNKT、NK、CD8 T 以及 CD4 T 细胞表面均表达。在小鼠中,CRTAM mRNA 在脾脏、大脑和睾丸中可检测到,在人体中则存在于脾脏、胸腺、肠道、淋巴结、肺、睾丸、卵巢和结肠中。研究发现,CRTAM+CD4 T 细胞产生的 IFN-γ 在 DSS 诱导的结肠炎小鼠模型中观察到的炎症过程中起到作用。在哮喘患者中检测到 CRTAM 存在于 CD4 和 CD8 T 细胞、中性粒细胞以及嗜碱性粒细胞的细胞膜上。此外,经抗原特异性刺激后,1 型糖尿病患者的 iNKT 细胞中 CRTAM 表达与 IFN-γ 产生之间存在明显关联。这些发现提示,CRTAM 表达细胞可能参与多种疾病相关的免疫病理学过程 [4-6]。
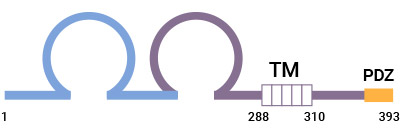
图 1. CRTAM 的结构 [1]
2. CRTAM 的配体是什么?
Nectin 样分子 2(Necl-2)最近被确认为 CRTAM 的结合伴侣。Necl-2 有很多别名,包括 TSLC1(tumor suppressor in lung cancer 1)、CADM1(cell adhesion molecule 1)、SynCAM1(synaptic cell adhesion molecule 1)、IgSF4(Immunoglobulin superfamily member 4)、RAI75。Necl-2 属于 Nectin 家族蛋白成员(Nectin1-4,Necl1-5)。Nectin 和 necl 分子之间通过形成同源或异源的二聚体介导细胞黏附,他们也可以和免疫分子如 DNAM-1(CD226),TIGIT,CD96,CRTAM 等互相作用。多个研究通过细胞粘附实验、NK 细胞与 CD8 细胞相关的功能实验鉴定出 CRTAM 的配体为 Nectin 家族蛋白 Necl-2 [5, 7-8]。
研究发现,CRTAM 与其配体 nectin-like 2(Necl2)的相互作用对小鼠 CD4 T 细胞产生 IL-17、IL-22 和 IFN-γ 的高效产生至关重要,并在 CD8 T 细胞和 NK 细胞的最佳细胞毒性中发挥作用。现有研究揭示,CRTAM 与 Necl-2 的结合产生不同效应,具体取决于淋巴细胞类型。对于 NK 或 CD8(+) T 细胞,它促进细胞毒性和 IFNγ 分泌,有利于免疫监视。相反,CRTAM/Necl-2(TSLC1)相互作用会触发激活的 Vγ9Vδ2 γδ T 细胞的细胞死亡,从而促进肿瘤免疫逃逸 [5, 7-9]。
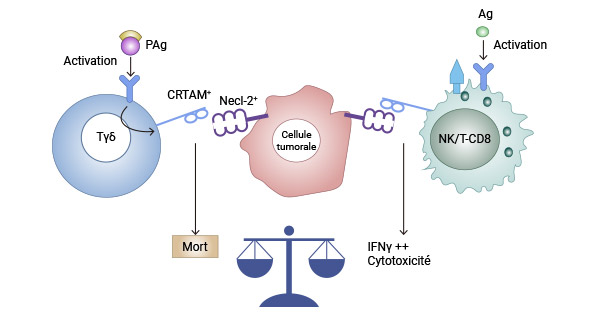
图 2. CRTAM 与 Necl-2 的结合产生不同效应 [9]
3. CRTAM 相关的调控机制
越来越多的研究表明,CRTAM 是一个多功能蛋白,在多种免疫过程中发挥作用。除了与 Necl-2 相互作用发挥免疫功能外,在小部分 CD4 T 细胞亚群中,CRTAM 可与蛋白 scrib 相互作用,调节 T 细胞的极性,调节 γ 干扰素和 IL-22 的分泌 [10-12]。此外,Zeb1 被发现是 CRTAM 的调控因子。Zeb1 抑制 CRTAM 启动子活性,调控人 T 细胞中的 CRTAM 水平 [13]。当 CRTAM 启动子中的特定元件突变时,可消除 Zeb1 介导的转录抑制 [13]。然而,AP-1 转录因子也被证明在人类 T CD8 淋巴细胞中调控 CRTAM 基因的表达 [14]。目前 CRTAM 介导的信号转导途径尚不十分清楚,多数研究提示 Necl-2 和 CRTAM 相互作用对免疫过程至关重要。
研究显示,CRTAM 能增强 NK 细胞对肿瘤细胞的杀伤能力。活化的 NK 细胞对转染了 Necl-2 的肿瘤细胞的杀伤效果更显著。体内实验表明,使用转染 Necl-2 的肿瘤细胞在裸鼠上进行实验时,裸鼠会迅速排斥肿瘤,预先清除裸鼠的 NK 细胞则明显抑制了排斥作用。体外实验表明,Necl-2 的肿瘤细胞活化 CD8+T 细胞时,CRTAM 表达增加,刺激后 CD8+T 细胞分泌 γ 干扰素。阻断 CRTAM-Necl-2 互动会抑制 γ 干扰素分泌。此外,Necl-2 的抗原递呈细胞可刺激 CD8+T 细胞分泌 IL-22,参与皮肤细胞的免疫调节。CRTAM-Necl-2 互动调节 CD8+T 细胞淋巴结停留。CRTAM 敲除小鼠对病毒免疫效果较弱,因其 CD8+T 细胞数量减少 [9-11]。
4. CRTAM 与疾病相关的研究
4.1 CRTAM 与肠道菌群研究
研究发现 CRTAM 存在于肠道内的 T 细胞中,参与调控 T 细胞在肠道的定位和维持,同时影响 Th17 细胞的分化和功能。CRTAM 的缺失影响了肠道菌群的组成,在平衡状态下,缺乏 CRTAM 的小鼠肠道中 Firmicutes 菌门的相对丰度增加,而 Tenericutes 菌门的相对丰度减少。研究使用野生型和缺乏 CRTAM 基因的小鼠,在感染或生理盐水灌胃后观察小鼠体重、粪便、肠道组织和肠道菌群的变化,以及血液和肠道淋巴结中 Th17 细胞的数量和功能。结果显示,CRTAM 促进了感染后 Th17 细胞的增殖和分化,增强了小鼠对肠道寄生虫的抵抗能力 [15]。
此外,CRTAM 还维持了肠道菌群的平衡,防止了感染后菌群失调和炎症反应的发生。与此相反,CRTAM 缺失的小鼠在感染后表现出更严重的体重下降、粪便稀释、肠道损伤和菌群失调,以及较低水平和功能的 Th17 细胞。因此,研究表明 CRTAM 是一种关键的免疫调节分子,其通过促进 Th17 细胞的成熟发挥保护作用,预防小鼠在肠道寄生虫感染中引起的菌群失调和炎症反应,为未来肠道寄生虫感染的治疗提供了新的靶点和策略 [15]。
4.2 CRTAM 与哮喘研究
研究发现过敏性哮喘患者的 CD4+CRTAM+和 CD8+CRTAM+T 淋巴细胞以及 CD177+CRTAM+中性粒细胞的百分比明显增加,而 IL5Rα+CRTAM+嗜酸性粒细胞的百分比无显著差异。因此,该研究推测 CRTAM 在 T 细胞、嗜酸性粒细胞和中性粒细胞的表达可能与过敏性哮喘的支气管炎症相关 [23]。另有研究指出维生素 D 水平与编码 CRTAM 基因的变异在哮喘发作风险中有显著的相互作用。在两个儿童哮喘队列(CAMP population 和 Costa Rican replication population)中进行了全基因组互作分析,揭示了三个与 CRTAM 相关的常见变异,其中 rs2272094 是一种非同义多态性,其在维生素 D 水平较低时与哮喘发作率的增加相关。进一步实验揭示了维生素 D 和 rs2272094 对 CRTAM 表达的影响。该研究结果在两个独立人群中得到了验证,揭示了维生素 D 与 CRTAM 调控的 CD8+和自然杀伤性 T 细胞在哮喘发作中的重要作用 [16]。
4.3 CRTAM 与糖尿病研究
CRTAM 在活化的 iNKT 细胞、CD8+T 细胞和少量 CD4+T 细胞表面得到表达,并与小鼠 CD4+T 细胞的促炎症表型相关。研究表明,受到 α-半乳糖脑苷刺激的 iNKT 细胞可在 18 小时内表达 CRTAM,这表明 iTCR 与 α-半乳糖脑苷的结合信号足以触发 CRTAM 的表达,而共刺激分子如 CD40、CD80 和 CD86 对 CRTAM 的表达并无影响。此外,研究人员还观察到 CRTAM 的表达与健康人和 I 型糖尿病患者的 iNKT 细胞产生干扰素-γ(IFN-γ)的能力有明显的关联。这表明 CRTAM 可能在人类 iNKT 细胞产生 IFN-γ 的过程中扮演重要角色,同时可作为识别这些炎症细胞的标志之一 [17]。
4.4 CRTAM 与癌症等疾病研究
在小鼠的肺癌模型中,研究者发现天花粉可通过调节肿瘤细胞中 TSLC1 和 CRTAM 的表达来发挥作用。进一步的实验发现,使用小干扰 RNA(siRNA)阻断 TSLC1 的表达可以显著消除天花粉对效应 T 细胞增殖和细胞因子分泌的影响。这表明天花粉通过促进 TSLC1 和 CRTAM 之间的相互作用来增强抗肿瘤免疫应答 [18];CRTAM 在免疫调节型三阴性乳腺癌(TNBC)中也具有重要作用。CRTAM 可增强免疫炎症反应并促进 CD8+T 细胞的浸润。其次,CRTAM 过表达能引发 STAT1 的磷酸化,并增加干扰素刺激基因的表达 [19]。此外,也有报道提示 CRTAM 与急性淋巴性白血病 [20]、自身免疫脱发 [21]、结核病 [22] 关联。总之,这些研究表明,CRTAM 参与调节免疫细胞的生物学行为和免疫应答,有望成为多种疾病的免疫研究新的靶点。
5. CRTAM 的临床研究前景
HBM-1054 作为针对 CRTAM 的临床药物,正在进行临床前的制剂开发和工艺优化,预计将在 2024 年提交临床试验申请。该药物由和铂医药(上海)有限责任公司开发,其临床适应症尚未公布,但可能涉及 CTRAM 高表达的肿瘤类型,比如肺癌、乳腺癌和胃癌等。当前对于 CRTAM 的临床药物开发和评价仍相对不足。因此,HBM-1054 的安全性和有效性需要更多的临床数据来证实。随着对 CRTAM 及其在肿瘤免疫治疗中作用的深入研究,更多的创新和合作是必要的,这将有助于扩大对 CRTAM 的理解,并为这一领域带来更多新型研究策略。
为鼎力协助科研和药企人员针对 CTRAM 在肠道、哮喘、糖尿病以及癌症等疾病中的临床应用研究,CUSABIO 推出 CTRAM 活性蛋白(CSB-MP005998 HU2; CSB-MP614820MO),助力您在 CTRAM 机制方面的研究或其潜在临床价值的探索。
华美 CUSABIO CTRAM 蛋白
● Recombinant Human Cytotoxic and regulatory T-cell molecule (CRTAM), partial (Active) Code: CSB-MP005998HU2
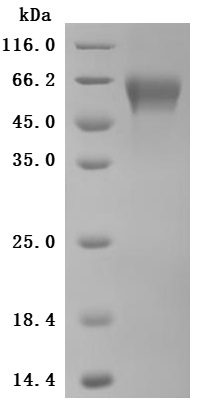
The high purity was greater than 95% as determined by SDS-PAGE.
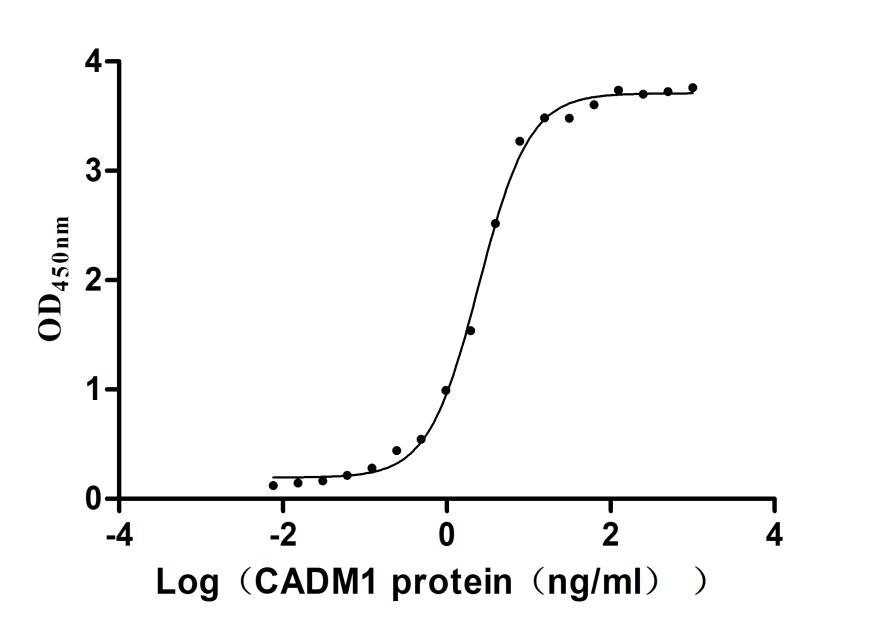
Immobilized Human CRTAM at 2μg/mL can bind Human CADM1(CSB-MP004425HUd9). The EC50 is 2.277-2.649 ng/mL.
● Recombinant Mouse Cytotoxic and regulatory T-cell molecule (Crtam), partial (Active) Code: CSB-MP614820MO
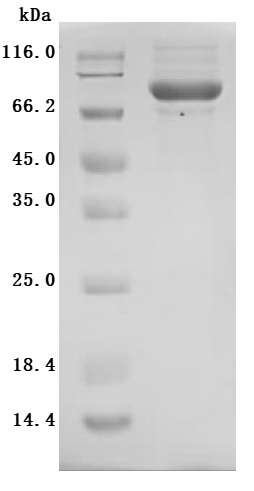
The high purity was greater than 90% as determined by SDS-PAGE.
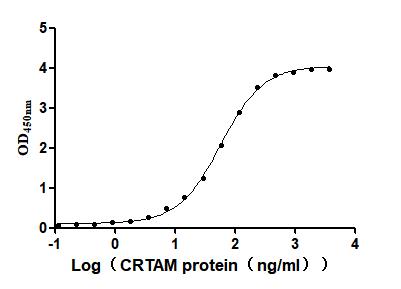
Immobilized Mouse Cadm1 (CSB-MP004425MO) at 2μg/mL can bind Mouse Crtam. The EC50 is 54.36 -61.77 ng/mL.
参考文献:
[1] Zhang Shuijun. Study on the molecular mechanism of interaction between NK cell receptors and Nectin/Necl family ligands [D]. China University of Science and Technology, 2014.
[2] Zhang, Shuijun, et al. "Competition of cell adhesion and immune recognition: Insights into the interaction between CRTAM and nectin-like 2." Structure 21.8 (2013): 1430-1439.
[3] Yeh, Jung-Hua, Sachdev S. Sidhu, and Andrew C. Chan. "Regulation of a late phase of T cell polarity and effector functions by Crtam." Cell 132.5 (2008): 846-859.
[4] Garay, Erika, et al. "CRTAM: A molecule involved in epithelial cell adhesion." Journal of cellular biochemistry 111.1 (2010): 111-122.
[5] Arase, Noriko, et al. "Heterotypic interaction of CRTAM with Necl2 induces cell adhesion on activated NK cells and CD8+ T cells." International immunology 17.9 (2005): 1227-1237.
[6] Galibert, Laurent, et al. "Nectin-like protein 2 defines a subset of T-cell zone dendritic cells and is a ligand for class-I-restricted T-cell-associated molecule*♦." Journal of Biological Chemistry 280.23 (2005): 21955-21964.
[7] Boles, Kent S., et al. "The tumor suppressor TSLC1/NECL-2 triggers NK-cell and CD8+ T-cell responses through the cell-surface receptor CRTAM." Blood 106.3 (2005): 779-786.
[8] Dessarthe, Benoît, et al. "CRTAM receptor engagement by Necl-2 on tumor cells triggers cell death of activated Vγ9Vδ2 T cells." The Journal of Immunology 190.9 (2013): 4868-4876.
[9] Nectins and nectin-like receptors DNAM-1 and CRTAM: New ways for tumor escape
[10] Leavy, Olive. "Polarity and CRTAM: a matter of timing." Nature Reviews Immunology 8.4 (2008): 246-246.
[11] Takeuchi, Arata, et al. "CRTAM determines the CD4+ cytotoxic T lymphocyte lineage." Journal of Experimental Medicine 213.1 (2016): 123-138.
[12] YEH, JUNG‐HUA, Sachdev S. Sidhu, and Andrew C. Chan. "Crtam regulates a late phase of T cell polarity and IFNγ/IL22 cytokine production." (2008): 384-384.
[13] Rojas-Marquez, C., et al. "CRTAM is negatively regulated by ZEB1 in T cells." Molecular Immunology 66.2 (2015): 290-298.
[14] Valle-Rios, Ricardo, et al. "Characterization of CRTAM gene promoter: AP-1 transcription factor control its expression in human T CD8 lymphocytes." Molecular immunology 46.16 (2009): 3379-3387.
[15] Perez-Lopez, Araceli, et al. "CRTAM shapes the gut microbiota and enhances the severity of infection." The Journal of Immunology 203.2 (2019): 532-543.
[16] Du, Rose, et al. "Genome-wide association study reveals class I MHC–restricted T cell–associated molecule gene (CRTAM) variants interact with vitamin D levels to affect asthma exacerbations." Journal of allergy and clinical immunology 129.2 (2012): 368-373.
[17] Beristain-Covarrubias, Nonantzin, et al. "Class I-restricted T cell-associated molecule is a marker for IFN-γ-producing iNKT cells in healthy subjects and patients with type 1 diabetes." Journal of Interferon & Cytokine Research 37.1 (2017): 39-49.
[18] Cai, Yuchan, et al. "Trichosanthin enhances anti-tumor immune response in a murine Lewis lung cancer model by boosting the interaction between TSLC1 and CRTAM." Cellular & molecular immunology 8.4 (2011): 359-367.
[19] Zheng, Shuyue, et al. "CRTAM promotes antitumor immune response in triple negative breast cancer by enhancing CD8+ T cell infiltration." (2023).
[20] Ramírez-Ramírez, Dalia, et al. "CRTAM+ NK cells endowed with suppressor properties arise in leukemic bone marrow." Journal of Leukocyte Biology 105.5 (2019): 999-1013.
[21] Giangreco, Adam, et al. "Epidermal Cadm1 expression promotes autoimmune alopecia via enhanced T cell adhesion and cytotoxicity." The Journal of Immunology 188.3 (2012): 1514-1522.
[22] Lai, Rachel PJ, et al. "Transcriptomic characterization of tuberculous sputum reveals a host Warburg effect and microbial cholesterol catabolism." Mbio 12.6 (2021): e01766-21.
[23] Ramirez-Velazquez, Carlos, et al. "Peripheral blood T cells and neutrophils from asthma patients express class-I MHC-restricted T cell-associated molecule." Allergy, Asthma & Clinical Immunology 10 (2014): 1-6.