- 移动端


武汉华美生物工程有限公司CUSABIO®品牌商
15 年
手机商铺
- NaN
- 0
- 0
- 2
- 2

推荐产品

公司新闻/正文
潜力新靶点ANGPTL4:连接代谢、炎症与组织重塑的关键分子
450 人阅读发布时间:2026-02-06 11:53
血管生成素样蛋白4(angiopoietin-like protein 4,ANGPTL4)是一种在多组织表达的分泌型糖蛋白,其生物学功能高度依赖组织来源、蛋白裂解状态及病理背景。研究表明,ANGPTL4通过N端结构域抑制脂蛋白脂肪酶(lipoprotein lipase,LPL)活性,调控脂肪酸在不同组织间的分配,而其C端片段则主要参与细胞黏附、炎症反应、血管通透性及组织重塑等非代谢过程。
近年来,大量证据显示ANGPTL4在代谢性心血管疾病、肿瘤、肾脏疾病、神经退行性疾病及纤维化过程中发挥情境依赖性的双重作用。本文系统梳理了ANGPTL4分子特征与核心信号通路,其在不同疾病中的作用机制、研究分歧及潜在转化价值,并总结当前靶向ANGPTL4的药物研发进展,为其作为生物标志物及治疗靶点的临床应用提供参考。
1. ANGPTL4的发现及基本生物学特征
2. ANGPTL4的核心分子机制与信号通路
3. ANGPTL4在代谢性疾病与心血管疾病中的作用
4. ANGPTL4在肿瘤生物学中的双重作用
5. ANGPTL4在肾脏、神经系统及纤维化疾病中的作用
6. 靶向ANGPTL4的药物研发进展
7. ANGPTL4研究工具
1. ANGPTL4的发现及基本生物学特征
ANGPTL4属于ANGPTL家族成员,最初被鉴定为脂肪组织分泌的脂肪因子,随后发现其在肝脏、肠道、肌肉、内皮细胞及多种肿瘤组织中均有表达。作为分泌蛋白,ANGPTL4可在胞外被蛋白水解为功能差异显著的N端与C端片段,其表达与裂解受营养状态、激素信号及炎症环境调控。
在人类内脏脂肪组织中,ANGPTL4在伴随葡萄糖代谢异常的肥胖个体中显著上调,并与脂肪组织炎症及脂肪细胞异质性相关,提示其可能参与代谢失衡的早期分子事件 [1]。体外研究显示,ANGPTL4可通过形成复合体或裂解产物调节其对LPL的抑制效应,这一特性为其在不同生理和病理状态下呈现差异化功能提供了分子基础 [2]。临床研究进一步将ANGPTL4与胰岛素抵抗、血管功能障碍及炎症状态相关联,奠定了其作为代谢—血管调节因子的研究基础 [3,4]。
2. ANGPTL4的核心分子机制与信号通路
从机制上看,ANGPTL4最明确、证据最充分的作用靶点是LPL。遗传学与功能研究一致表明,ANGPTL4通过抑制LPL活性限制富含甘油三酯脂蛋白的水解,从而调控脂肪酸进入脂肪组织、肌肉和肝脏的速率 [7,9]。这一作用在脂肪组织中尤为关键,其局部抑制或缺失可显著改善全身脂质分配与胰岛素敏感性 [7]。
除LPL外,ANGPTL4还通过与整合素、BMP/TGF-β信号组件及AKT/MTOR等通路发生互作,参与细胞存活、炎症反应及组织重塑 [13,16]。这些通路并非在所有组织中同时激活,而是高度依赖细胞类型、蛋白片段形式及微环境因素,这种多层级调控构成了ANGPTL4功能异质性的分子基础。
3. ANGPTL4在代谢性疾病与心血管疾病中的作用
在脂质代谢调控中,ANGPTL4通过抑制LPL改变组织对VLDL等富含甘油三酯脂蛋白的脂肪酸摄取,是调控血浆甘油三酯水平的重要因子。肝脏作为系统性脂质代谢枢纽,其分泌的ANGPTL4对外周脂蛋白代谢具有显著影响。
肝靶向反义寡核苷酸(ASO)沉默Angptl4在APOE*3-Leiden.CETP小鼠中显著降低血浆甘油三酯和胆固醇水平,并明显缩小动脉粥样硬化斑块面积,同时在非人灵长类动物中表现出较好的短期耐受性,提示肝脏特异性干预在疗效与安全性之间具备优势 [19]。与此一致,肝细胞特异性Angptl4缺失模型通过增强肝脂蛋白酯酶活性,加速残余脂蛋白清除,从而改善肥胖、胰岛素抵抗和动脉粥样化表型 [20]。
人群遗传学证据进一步支持ANGPTL4在血脂调控中的因果作用。E40K等功能缺失变异与较低甘油三酯水平和更有利的代谢表型相关,并在肥胖人群中表现出一定的保护效应 [21]。然而,遗传变异的终身效应与药物干预的短期或中期作用并不完全等同,这一差异在解读转化潜力时需要谨慎权衡。
此外,ANGPTL4还被纳入高血压和代谢综合征的分子网络中。特定位点SNP与腰围、血压及代谢综合征风险相关 [22],而高血压患者中循环及脂肪组织ANGPTL4升高的观察结果提示其可能参与血管张力调控和内皮功能改变 [23]。与此同时,肠道微生物通过调控肠道ANGPTL4表达影响脂质吸收和能量平衡,进一步凸显了其在“肝—肠—外周组织”轴中的系统性作用 [24,25]。
4. ANGPTL4在肿瘤生物学中的双重作用
ANGPTL4在肿瘤中的作用呈现高度情境依赖性,其既可作为促肿瘤因子,也可在特定背景下发挥抑癌效应。
在部分肿瘤模型中,ANGPTL4表现出抑癌特性。例如,在宫颈癌中,ANGPTL4过表达可抑制肿瘤细胞增殖、迁移和侵袭,并增强顺铂诱导的凋亡反应,而敲低则产生相反效应,提示其可能作为化疗增敏因子。然而,这些结论主要基于体外模型,尚缺乏体内和临床样本层面的验证。
在清晰细胞肾细胞癌中,ANGPTL4低表达与不良预后相关,其N端片段可通过抑制溶酶体酸性脂肪酶(LAL)限制脂质代谢驱动的肿瘤生长,提示ANGPTL4缺失状态下肿瘤可能更依赖脂质代谢通路。
与上述抑癌效应相对,ANGPTL4在肥胖相关肿瘤微环境中常表现为促癌因子。在三阴性乳腺癌中,脂肪组织分泌因子通过PPARα-ANGPTL4轴重编程肿瘤脂质代谢,并通过FAK信号增强侵袭和迁移能力 [5]。此外,在肺腺癌中,ANGPTL4通过抑制焦亡和凋亡通路,促进EGFR抑制剂耐药,揭示其在治疗耐受形成中的潜在作用 [8]。
值得注意的是,ANGPTL4裂解片段在肿瘤中的功能存在显著差异。研究表明,C端片段可促进肿瘤生长和转移,而N端片段则抑制转移并降低血管生成,这一现象强调蛋白加工状态是决定其肿瘤学效应的关键变量。
5. ANGPTL4在肾脏、神经系统及纤维化疾病中的作用
在肾脏疾病中,ANGPTL4被证明参与脂质沉积、细胞衰老及纤维化进程。肥胖和衰老小鼠肾组织中ANGPTL4显著上调,其过表达可促进肾小管上皮细胞脂质积累和衰老表型,而基因抑制则具有保护作用 [10]。在糖尿病肾病中,ANGPTL4通过与Integrin β1互作并激活STING通路,促进炎症反应和上皮-间质转化,从而加速肾间质纤维化 [11]。
在中枢神经系统,ANGPTL4被认为连接代谢异常与微胶质细胞功能失调。在阿尔茨海默病模型中,其上调与脂滴富集型微胶质细胞表型、氧化应激和炎症增强密切相关,提示其可能介导肥胖与神经退行性疾病之间的病理联系 [12]。
在血管生成和纤维化疾病中,ANGPTL4通过促进内皮-间质转化(EndMT)参与脉络膜新生血管和视网膜下纤维化形成,并可能影响抗VEGF治疗反应 [17,18]。在肺纤维化模型中,ANGPTL4通过调控糖酵解重编程驱动间皮-间质转化,提示其作为代谢-表型转换节点的潜在价值 [6]。
6. 靶向ANGPTL4的药物研发进展
ANGPTL4正成为代谢与炎症相关疾病领域的一个新兴药物靶点。目前,围绕该靶点已涌现出多种技术路线的在研药物,涉及抗体、寡核苷酸及mRNA等不同类型,主要聚焦于脂代谢异常、心血管疾病及呼吸系统感染等适应症。全球多家研发机构参与推进,部分项目已进入临床中期试验,整体研发进程活跃。部分研发管线整理如下表:
| 药物 | 作用机制 | 药物类型 | 在研适应症 | 在研机构 | 最高研发阶段 |
|---|---|---|---|---|---|
| MAR-001 | ANGPTL4抑制剂 | 单克隆抗体 | 高甘油三酯血症 | 动脉粥样硬化 | Marea Therapeutics Pty Ltd. | 临床2期 |
| Lipisense | ANGPTL4抑制剂 | ASO | 高甘油三酯血症 | 代谢综合征 | 家族性乳糜微粒血症综合征 | Apotek Produktion & Laboratorier AB | Secarna Pharmaceuticals Gmbh & Co. | 成都先衍生物技术有限公司 | Lipigon Pharmaceuticals AB | 临床2期 |
| CR-064 | ANGPTL4抑制剂 | Ang1抑制剂 | 单克隆抗体 | 肾细胞癌 | CuraGen Corp. | 临床前 |
| ARDS treatment (Lipigon) | ANGPTL4抑制剂 | ASO | 急性呼吸窘迫综合征 | Lipigon Pharmaceuticals AB | 临床前 |
| P4 CAP (Lipigon Pharmaceuticals) | ANGPTL4抑制剂 | ASO | 社区获得性肺炎 | Lipigon Pharmaceuticals AB | 临床前 |
| WO2023044458 | - | 寡核苷酸 | 消化系统疾病 | 内分泌与代谢疾病 | 纤维化 | Alnylam Pharmaceuticals, Inc. | 药物发现 |
| CN118853662 | - | mRNA | 心血管疾病 | 消化系统疾病 | 内分泌与代谢疾病 | 罕见病 | 成都先衍生物技术有限公司 | 药物发现 |
(数据截止到2026年1月28日,来源于synapse)
7. ANGPTL4研究工具
ANGPTL4是连接脂质代谢、炎症反应与组织重塑的关键分子,其生物学效应高度依赖组织来源、蛋白片段形式及病理背景。尽管现有研究已揭示其在多系统疾病中的重要作用,但功能异质性和物种差异仍是临床转化的主要挑战。华美生物提供ANGPTL4重组蛋白、抗体、ELISA检测试剂盒相关产品,助力您的基础科研或药物开发:
● ANGPTL4重组蛋白
Recombinant Human Angiopoietin-related protein 4 (ANGPTL4), partial
CSB-EP866314HU1
Recombinant Mouse Angiopoietin-related protein 4 (Angptl4)
CSB-MP4186MO
● ANGPTL4抗体
ANGPTL4 Antibody; CSB-PA866314ESR2HU
ANGPTL4 Antibody
CSB-PA866314EA01HU
ANGPTL4 (MAR-001 Biosimilar) Recombinant Monoclonal Antibody
CSB-RA866314MB1HU
● ANGPTL4 ELISA试剂盒
Human Angiopoietin-related protein 4(ANGPTL4) ELISA kit
CSB-EL001712HU
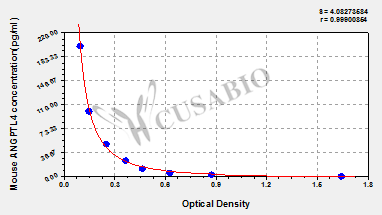
Mouse Angiopoietin-related protein 4(ANGPTL4) ELISA kit
CSB-EL001712MO
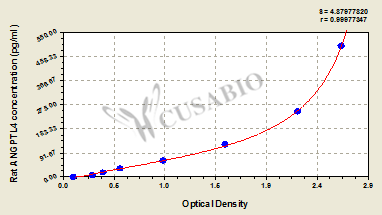
Rat Angiopoietin-related protein 4(ANGPTL4) ELISA kit
CSB-EL001712RA
[1] Ilaria Barchetta, Caterina Chiappetta, Valentina Ceccarelli, Flavia Agata Cimini, Laura Bertoccini, Melania Gaggini, Claudio Di Cristofano, Gianfranço Silecchia, Andrea Lenzi, Frida Leonetti, Marco Giorgio Baroni, Amalia Gastaldelli, Maria Gisella Cavallo.(2020). Angiopoietin-Like Protein 4 Overexpression in Visceral Adipose Tissue from Obese Subjects with Impaired Glucose Metabolism and Relationship with Lipoprotein Lipase.
[2] Günther Silbernagel, Yan Q Chen, Hongxia Li, Deven Lemen, Yi Wen, Eugene Y Zhen, Martin Rief, Marcus E Kleber, Graciela E Delgado, Mark A Sarzynski, Yue-Wei Qian, Boerge Schmidt, Raimund Erbel, Ulrike S Trampisch, Angela P Moissl, Henrik Rudolf, Heribert Schunkert, Andreas Stang, Winfried März, Hans J Trampisch, Hubert Scharnagl, Robert J Konrad.(2025). Associations of Circulating ANGPTL3, C-Terminal Domain-Containing ANGPTL4, and ANGPTL3/8 and ANGPTL4/8 Complexes with LPL Activity, Diabetes, Inflammation, and Cardiovascular Mortality.
[3] Haohua Wang, Wanying Luo, Min Lin, Xiaojing Li, Guangda Xiang, Silvia d Triganti.(2021). Plasma asprosin, CCDC80 and ANGPTL4 levels are associated with metabolic and cardiovascular risk in patients with inflammatory bowel disease.
[4] Francesca Schinzari, Giuseppina Vizioli, Umberto Campia, Manfredi Tesauro, Carmine Cardillo.(2021). Variable Changes of Circulating ANGPTL3 and ANGPTL4 in Different Obese Phenotypes: Relationship with Vasodilator Dysfunction.
[5] Christina Blücher, Sabine Iberl, Nancy Schwagarus, Silvana Müller, Gerhard Liebisch, Marcus Höring, Maria Soledad Hidrobo, Josef Ecker, Nick Spindler, Arne Dietrich, Ralph Burkhardt, Sonja C. Stadler.(2020). Secreted Factors from Adipose Tissue Reprogram Tumor Lipid Metabolism and Induce Motility by Modulating PPARα/ANGPTL4 and FAK.
[6] Yuechong Xia, Fang Zhou, Hongyan Hui, Liping Dai, Songyun Ouyang.(2024). ANGPTL4 mediated mesothelial-mesenchymal transition in pulmonary fibrosis: a potential therapeutic target.
[7] Binod Aryal, Abhishek K. Singh, Xinbo Zhang, Luis Varela, Noemí Rotllán, Leigh Goedeke, Balkrishna Chaube, João Paulo Camporez, Daniel F. Vatner, Tamas L. Horváth, Gerald I. Shulman, Yajaira Suárez, Carlos Fernández‐Hernando.(2018). Absence of ANGPTL4 in adipose tissue improves glucose tolerance and attenuates atherogenesis.
[8] Yue Fang, Xuan Li, Hao Cheng, Lu Zhang, Jiqing Hao.(2022). ANGPTL4 Regulates Lung Adenocarcinoma Pyroptosis and Apoptosis via NLRP3\ASC\Caspase 8 Signaling Pathway to Promote Resistance to Gefitinib.
[9] Fredrik Landfors, Elin Chorell, Sander Kersten.(2022). Genetic Mimicry Analysis Reveals the Specific Lipases Targeted by the ANGPTL3-ANGPTL8 Complex and ANGPTL4.
[10] Xiaojun Wang, Hung-Chen Chang, Xuchao Gu, Wanlin Han, Shihang Mao, Lili Lu, Shuai Jiang, Haiyong Ding, Shisheng Han, Xinkai Qu, Zhijun Bao.(2024). Renal lipid accumulation and aging linked to tubular cells injury via ANGPTL4.
[11] Swayam Prakash Srivastava, Han Zhou, Rachel Shenoi, Myshal Morris, Begoña Lainez, Leigh Goedeke, Barani Kumar Rajendran, Ocean Setia, Binod Aryal, Keizo Kanasaki, Daisuke Koya, Ken Inoki, Alan Dardik, Thomas Bell, Carlos Fernández‐Hernando, Gerald I. Shulman, Julie E. Goodwin.(2024). Renal Angptl4 is a key fibrogenic molecule in progressive diabetic kidney disease.
[12] Nan Li, Xiaojun Wang, Ruilang Lin, Fuxia Yang, Hung-Chen Chang, Xuchao Gu, Jun Shu, Guidong Liu, Yongfu Yu, Wenshi Wei, Zhijun Bao.(2024). ANGPTL4-mediated microglial lipid droplet accumulation: Bridging Alzheimer's disease and obesity.
[13] Yun Bai, Guanghua Cui, Xiaoke Sun, Meiqi Wei, Yanying Liu, Jialu Guo, Yu Yang.(2024). ANGPTL4 Stabilizes Bone Morphogenetic Protein 7 Through Deubiquitination and Promotes HCC Proliferation via the SMAD/MAPK Pathway.
[14] Xiujin Shen, Haibing Wang, Chunhua Weng, Yongchun He, Xue Shao, Jingyun Le, Hui Chen, Qixia Shen, Jianghua Chen, Hong Jiang.(2025). Angiopoietin-like 4 exacerbates renal tubular epithelial cell pyroptosis in acute kidney injury via integrin β5/FAK signaling pathway.
[15] Tin May Aung, Atit Silsirivanit, Apinya Jusakul, Waraporn Chan-On, Tanakorn Proungvitaya, Sittiruk Roytrakul, Siriporn Proungvitaya.(2022). Prediction of Angiopoietin-like Protein 4-related Signaling Pathways in Cholangiocarcinoma Cells.
[16] Xiao Song, Wang Nai‐dong, Jinxiang Yan, Long Tian, Xiurong Lu, Gao Hong, Yan Jie‐Cheng, Zhang Fei.(2022). ANGPTL4 regulate glutamine metabolism and fatty acid oxidation in nonsmall cell lung cancer cells.
[17] Jia Chen, Ying Yang, Shu Su, Shenglai Zhang, Ju Huang, Hong Chen, Xiaowei Yang, Aimin Sang.(2024). ANGPTL4 promotes choroidal neovascularization and subretinal fibrosis through the endothelial‒mesenchymal transition.
[18] Deepti Sharma, Evan Lau, Yu Qin, Kathleen Jee, Murilo Rodrigues, Chuanyu Guo, Aumreetam Dinabandhu, Emma McIntyre, Shaima Salman, Yousang Hwang, Ala Moshiri, Gregg L Semenza, Silvia Montaner, Akrit Sodhi.(2024). VEGF inhibition increases expression of HIF-regulated angiogenic genes by the RPE limiting the response of wet AMD eyes to aflibercept.
[19] Melanie Modder, Wietse In Het Panhuis, Mohan Li, Salwa Afkir, Alexandra L Dorn, Amanda C M Pronk, Trea C M Streefland, Reshma A Lalai, Stefan Pierrou, Stefan K Nilsson, Gunilla Olivecrona, Sander Kooijman, Patrick C N Rensen, Milena Schönke.(2024). Liver-targeted Angptl4 silencing by antisense oligonucleotide treatment attenuates hyperlipidaemia and atherosclerosis development in APOE*3-Leiden.CETP mice.
[20] Abhishek K. Singh, Balkrishna Chaube, Xinbo Zhang, Jonathan Sun, Kathryn M. Citrin, Alberto Canfrán‐Duque, Binod Aryal, Noemí Rotllán, Luis Varela, Richard Lee, Tamas L. Horváth, Nathan L. Price, Yajaira Suárez, Carlos Fernández‐Hernando.(2021). Hepatocyte-specific suppression of ANGPTL4 improves obesity-associated diabetes and mitigates atherosclerosis in mice.
[21] Diego Bailetti, Laura Bertoccini, Rosellina Margherita Mancina, Ilaria Barchetta, Danila Capoccia, Efisio Cossu, Arturo Pujia, Andrea Lenzi, Frida Leonetti, Maria Gisella Cavallo, Stefano Romeo, Marco Giorgio Baroni.(2018). ANGPTL4 gene E40K variation protects against obesity‐associated dyslipidemia in participants with obesity.
[22] Zhoujie Tong, Jie Peng, Hongtao Lan, Wenwen Sai, Yulin Li, Jiaying Xie, Yanmin Tan, Wei Zhang, Ming Zhong, Zhihao Wang.(2021). Cross-talk between ANGPTL4 gene SNP Rs1044250 and weight management is a risk factor of metabolic syndrome.
[23] Mohamed Abu‐Farha, Preethi Cherian, Mohamed G. Qaddoumi, Irina Al‐Khairi, Sriraman Devarajan, Muath Alanbaei, Jehad Abubaker.(2018). Increased plasma and adipose tissue levels of ANGPTL8/Betatrophin and ANGPTL4 in people with hypertension.
[24] Jing Guo, Mengyuan Zhang, He Wang, Na Li, Zongliang Lu, Long Li, Suocheng Hui, Hongxia Xu.(2022). Gut microbiota and short chain fatty acids partially mediate the beneficial effects of inulin on metabolic disorders in obeseob/obmice.
[25] Sang‐Hyun Cho, Yong‐Joon Cho, Joo‐Hong Park.(2021). The human symbiont Bacteroides thetaiotaomicron promotes diet-induced obesity by regulating host lipid metabolism.