- 移动端


武汉华美生物工程有限公司CUSABIO®品牌商
15 年
手机商铺
- NaN
- 0
- 0
- 2
- 2

推荐产品

公司新闻/正文
DLL3靶点新突破:再鼎医药公布ZL-1310早期临床数据!
2331 人阅读发布时间:2024-10-30 13:48
DLL3靶点新突破:再鼎医药公布ZL-1310早期临床数据!
2024年10月24日,再鼎医药在EORTC-NCI-AACR(ENA)会议上宣布,其开发的ZL-1310,一种靶向DLL3的抗体药物偶联物(ADC),在1期临床试验中展现出显著的疗效和良好的安全性和耐受性。该试验设计为单臂、盲法、国际多中心试验,受试者为年龄在18岁以上的成年男性和女性,患有经组织学或细胞学证实的转移性或广泛期小细胞肺癌。数据显示,ZL-1310在广泛期小细胞肺癌患者中达到了74%的客观缓解率(ORR),表明其具有显著的抗肿瘤活性 [1]。与此同时,宜联生物与安进达成合作协议,共同开展靶向DLL3和CD3的双特异性药物tarlatamab的临床试验,用于治疗小细胞肺癌。Tarlatamab已于2024年5月获得FDA加速批准上市,它能够激活患者自身的T细胞,特异性地杀伤表达DLL3的肿瘤细胞 [2]。这些前沿进展标志着DLL3作为小细胞肺癌治疗的重要靶点,正逐步实现从实验室到临床的转化,为患者带来更多的治疗选择和希望。
1、DLL3的结构和功能是什么?
δ样配体(Delta-Like Ligand 3,DLL3)为一种附着在细胞表面的单次跨膜蛋白,属于Notch配体家族中的一员。人DLL3基因定位于染色体19q13,其开放阅读框长度约为1800 bp [3]。人DLL3蛋白由619个氨基酸组成,完整结构包含1个DSL结构域、1个胞内结构域和6个表皮生长因子样结构域 (图1)[4, 5]。胞外结构域N端的DSL基因序列在配体家族中高度保守,是与Notch受体结合所必须的功能结构域。DLL3胞内结构域较短,其功能尚不清楚。研究发现DLL3在SCLC和其他神经内分泌肿瘤中高表达,而在正常组织中很少表达,这为靶向治疗提供了潜力 [6]。近年来,越来越多的研究提示,DLL3可与不同的Notch受体结合,参与到复杂的肿瘤调控过程中,既可表现出促癌作用,也可表现出抑癌作用。
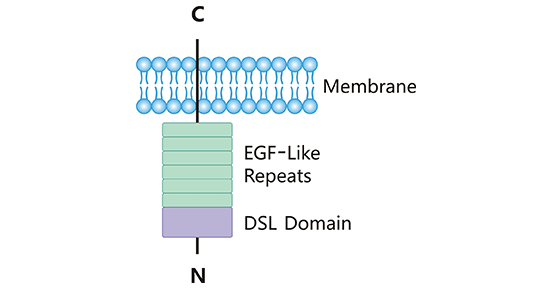
图1. DLL3结构示意图
*图片来源于Frontiers in immunology 出版物 [4]
2、配体DLL3的受体有哪些?
配体DLL3通过与Notch受体结合,直接或间接地发挥作用生物学功能。Notch受体有4种(Notch 1-4),为一类保守的单次跨膜蛋白,受体分子量约为300 kDa [7]。Notch受体含胞外区、跨膜区和胞内区3部分。如图2所示,其Notch受体的胞外段(N端)由数量不等的EGF样重复片段和一个近膜区的负向调节区域(NRR)构成,而NRR是由3个Lin12/Notch重复片段(LNR)和1个异二聚化区域(HD)组成;胞内段通常包含一段蛋白结合RPBJ相关分子(RAM)区域、7个锚蛋白重复片段、1个转录激活区域(TAD)和1个PEST(富含脯氨酸、谷氨酸、丝氨酸和苏氨酸)降解决定子区域 [4]。
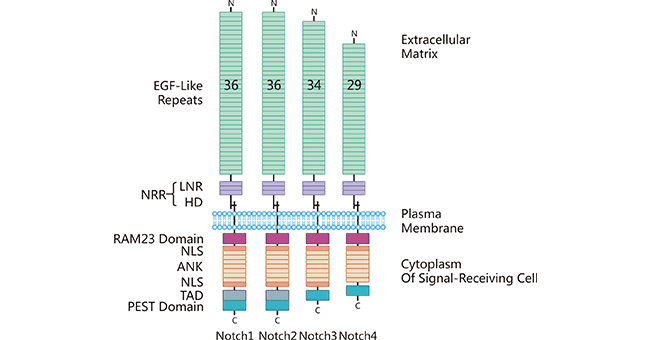
图2. 受体Notch1-4结构
*图片来源于Frontiers in immunology 出版物 [4]
不同于Notch其它配体,目前的研究表明DLL3是一种抑制性Notch配体。配体DLL3和Notch受体结合,具有Notch通路的抑制作用 [8]。在SCLC中,促进DLL3表达,DLL3与Notch1受体结合,将抑制Notch信号活化,Notch信号靶基因HES1、HEY1的表达下调,对肿瘤的抑制解除,促进SCLC的发生发展 [9];DLL3/Notch2可以增加细胞周期蛋白CyclinD1和CyclinD3的表达,提示DLL3/Notch2可能通过上调CyclinD1和CyclinD3蛋白促进SCLC细胞的增殖 [10]。此外DLL3和Notch受体结合还与其它肿瘤密切相关,比如,DLL3/Notch2参与垂体腺瘤的增殖和侵袭的调节 [11];DLL3/Notch2/Notch4对黑色素瘤细胞生存和生长至关重要 [12];DLL3/Notch2/Notch3介导了卵巢癌细胞的增殖和分化,并且与较差的生存期相关 [13]。总体而言,DLL3可与不同的Notch受体结合,在细胞增殖、分化及凋亡中发挥着多种功能,然而其涉及的细胞分子机制仍未具体阐明。
3、DLL3相关的信号通路有哪些?
前文提到,DLL3是一种抑制性Notch配体,大量研究发现,配体DLL3通过和Notch受体结合,抑制Notch信号,影响相邻细胞之间的通讯,进而调控细胞发育,其具体机制尚不明确。除了Notch信号通路,DLL3也在其他信号通路中发挥作用,DLL3通过抑制Notch信号通路,激活脂酰肌醇-3-激酶/丝氨酸-苏氨酸蛋白激B(phosphoinositol-3-kinase/serine-threonine protein kinase B,P13K/Akt)信号传导通路。DLL3表达上调时,配体Wnt-1和Wnt-4以及Wnt通路的下游靶基因Axin-2和Lef-1表达上调,提示DLL3参与诱导Wnt信号途径的激活。此外,还有研究证实DLL3通过调节Nrarp的循环表达来调节Notch/Wnt信号通路 [14]。
由此可见,DLL3参与到多个信号通路中的调节过程,涉及一系列基因的激活、表达以及调控等作用。在肿瘤的发生发展中,DLL3发挥着促癌或抑癌的双向调节功能。因此,进一步研究DLL3在各肿瘤中的表达和作用机制,将对肿瘤的发病机制、治疗和预防等具有重要的意义。
4、DLL3在肿瘤疾病中的作用
根据肿瘤类型和细胞生长环境的不同,DLL3的激活可以发挥促癌或抑癌作用。有报道,DLL3在小细胞肺癌 [15, 16]、乳腺癌 [17]、垂体瘤 [11]、急性髓系白血病 [18]中有促癌作用,但在肝癌 [19]、神经胶质瘤 [20]和恶性胶质瘤 [21]中却发挥了抑癌作用。
在小细胞肺癌中,DLL3在超过80%患者中高表达,并且在肿瘤的细胞膜和细胞质中都高表达;但是,在正常组织中少量表达或不表达。临床研究表明,SCLC中的DLL3高表达与患者的生存期呈现负相关,即DLL3表达量越高,患者的生存期越低 [22]。
在原发性肝癌中,有研究提示,DLL3的表达被乙肝病毒诱导的DNA甲基化和组蛋白乙酰化所抑制。抑制组蛋白去乙酰化酶的抑制剂可以使DLL3在HCC中重新表达。重表达的DLL3可以抑制HCC细胞的生长并诱导细胞凋亡。因此,在原发性肝癌中,DLL3可以抑制癌细胞的生长 [23]。
更多研究显示DLL3在多种癌症中异常表达,发挥不同作用。比如,DLL3在异柠檬酸脱氢酶IDH突变的神经胶质瘤细胞中表达,特别是在1p/19q缺失的神经胶质瘤细胞中高表达 [24, 25];在胰腺癌中,激活的DLL3可以刺激Notch信号从而促进癌细胞的生长 [26];在黑色素瘤中,DLL3/MAPK通路可以促进黑色素瘤细胞的增殖和迁移 [27, 28];在子宫内膜瘤中,高表达的DLL3与较差的生存期和较差的无进展生存期相关 [29]。
5、靶向DLL3的研发药物及临床意义
DLL3已经成为临床药物研究中的热门靶点,目前有超过50种药物正在积极研发中,例如AMG 757、BI 764532和BHP-01。这些药物采用多种作用机制,如AMG 757通过可裂解连接子与喜树碱衍生物结合,BHP-01则利用修饰的嵌合抗原受体T细胞技术。它们主要针对肿瘤领域,覆盖多种肿瘤类型,研发阶段从临床前到已在美国获得加速批准(如AMG 757)或快速通道(如BI 764532)不等。药物类型多样,包括抗体药物、CAR-T、小分子药物疗法。众多机构如勃林格殷格翰、安进、百济神州、泽璟制药、齐鲁制药等参与了研发,复旦大学和Compugen公司也参与了相关研究,Compugen与吉利德签订了合作协议。这表明DLL3药物研发在全球范围内非常活跃,其治疗潜力得到了广泛认可。
特别是在小细胞肺癌(SCLC)领域,再鼎医药的ZL-1310作为国内前沿DLL3 ADC,在Ia期临床研究中展现了卓越的客观缓解率和良好的安全性,这标志着DLL3靶向药物在克服初期开发障碍后取得了重大进展。ZL-1310的成功不仅为DLL3作为SCLC的“定制”靶点提供了有力证据,也将推动DLL3赛道的发展,吸引更多制药企业的关注,促进全球制药行业在SCLC治疗上的创新和进步。随着ZL-1310及其他DLL3靶向药物如AMG 757、BI 764532和BHP-01的研发进展,预计这一领域将取得更多突破,为患者提供更多选择。
6、DLL3科研产品服务
为鼎力协助各药企针对DLL3靶点在肿瘤中的研发工作,尤其是在小细胞肺癌SCLC中,大量的基础研究和临床研究证据表明DLL3是治疗SCLC的一种极具潜力的干预靶点,CUSABIO已推出DLL3系列产品,种属多样、标签多样,能够充分满足不同种属交叉实验及各类实验需求,为科研人员提供优质的产品,助力生命科学领域的研究与发展。
DLL3蛋白
● Recombinant Human Delta-like protein 3 (DLL3), partial (Active), His-tagged (Code: CSB-MP882142HU3d7)
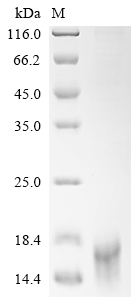
Purity ≥ 95% validated by SDS-PAGE
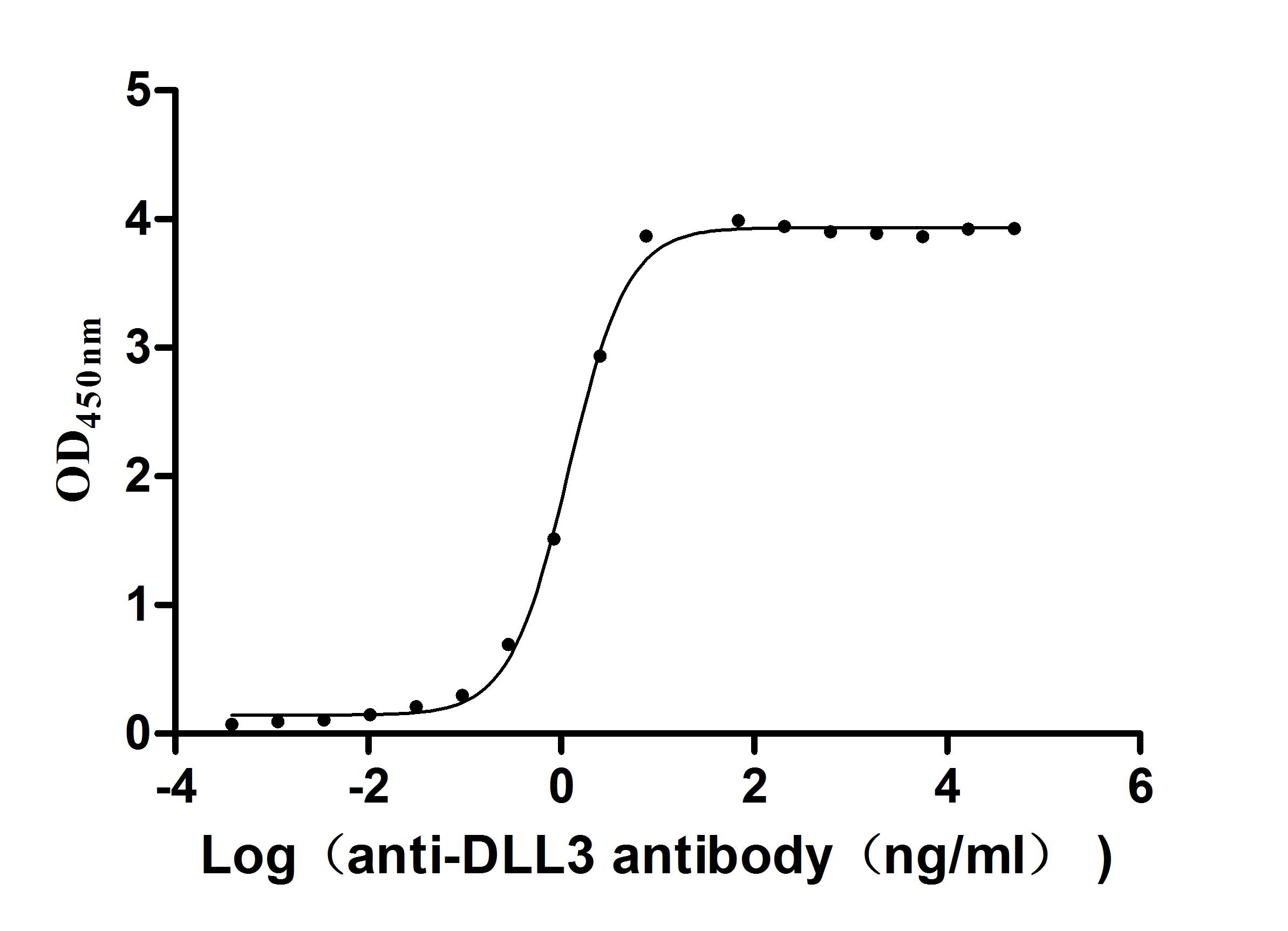
Activity validated by Functional ELISA: Measured by its binding ability in a functional ELISA. Immobilized Human DLL3 at 2 μg/ml can bind Anti-DLL3 recombinant antibody (CSB-RA882142MA2HU). The EC50 is 1.107-1.282 ng/mL.
● Recombinant Human Delta-like protein 3 (DLL3), partial (Active),Fc-tagged (Code: CSB-MP882142HU2)
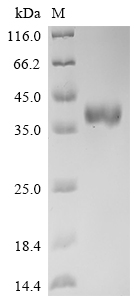
Purity ≥ 95% validated by SDS-PAGE
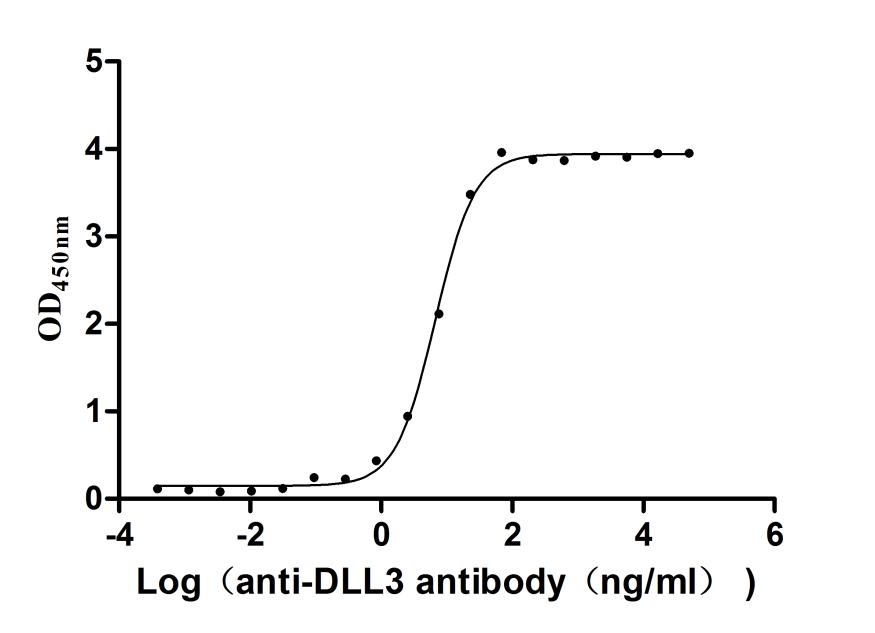
Activity validated by Functional ELISA: Measured by its binding ability in a functional ELISA. Immobilized Human DLL3 at 1 μg/ml can bind Anti-DLL3 recombinant antibody (CSB-RA882142MA2HU). The EC50 is 6.211-7.209 ng/mL.
● Recombinant Macaca fascicularis Delta-like protein 3 (DLL3), partial (Active),His-tagged (Code: CSB-MP3536MOV)
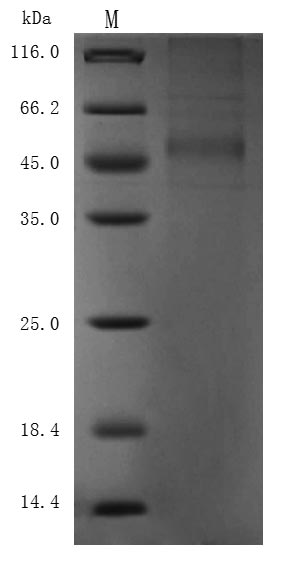
Purity ≥ 85% validated by SDS-PAGE
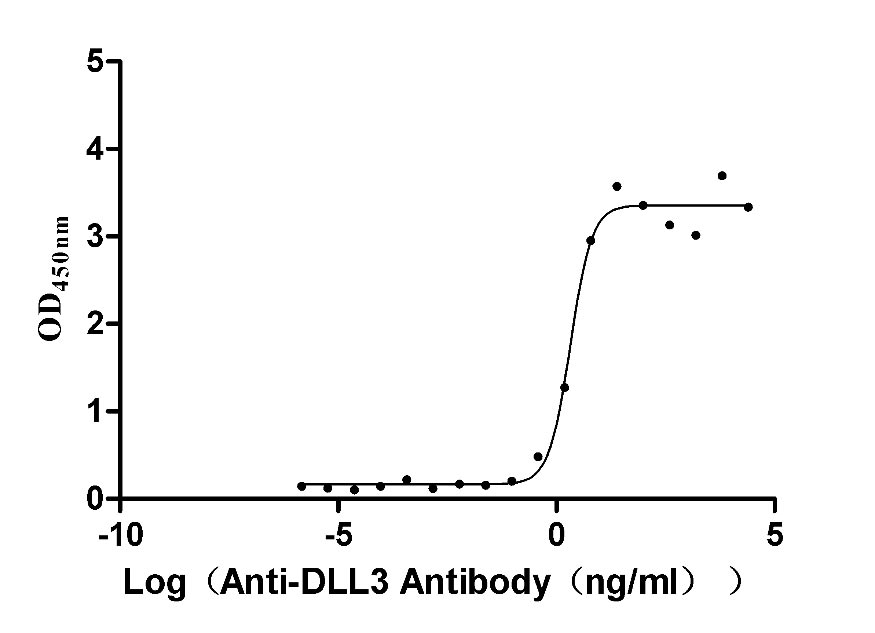
Activity validated by Functional ELISA: Measured by its binding ability in a functional ELISA. Immobilized DLL3 at 2 μg/ml can bind Anti-DLL3 Recombinant Antibody (CSB-RA882142A1HU). The EC50 is 1.625-2.702 ng/mL.
DLL3抗体
DLL3 Recombinant Monoclonal Antibody (ELISA, FC) (CSB-CSB-RA882142A1HU)
DLL3试剂盒
Human DLL3 ELISA Kit (CSB-EL006948HU)
参考文献:
[1] xueqiu.com/4797094524/309486542
[2] 163.com/dy/article/JDVKQLMO0552Q7S2.html?spss=dy_author
[3] Bulman, Michael P., et al. "Mutations in the human delta homologue, DLL3, cause axial skeletal defects in spondylocostal dysostosis." Nature genetics 24.4 (2000): 438-441.
[4] Steinbuck, Martin Peter, and Susan Winandy. "A review of notch processing with new insights into ligand-independent notch signaling in T-cells." Frontiers in immunology 9 (2018): 1230.
[5] Hu, Bingxin, et al. "Over-expression of human Notch ligand Delta-like 3 promotes proliferation of human gastric cancer cells in vitro." Nan Fang yi ke da xue xue bao= Journal of Southern Medical University 38.1 (2018): 14-19.
[6] Saunders, Laura R., et al. "A DLL3-targeted antibody-drug conjugate eradicates high-grade pulmonary neuroendocrine tumor-initiating cells in vivo." Science translational medicine 7.302 (2015): 302ra136-302ra136.
[7] Stanley, Pamela. "Regulation of Notch signaling by glycosylation." Current opinion in structural biology 17.5 (2007): 530-535.
[8] Chapman, Gavin, et al. "Notch inhibition by the ligand DELTA-LIKE 3 defines the mechanism of abnormal vertebral segmentation in spondylocostal dysostosis." Human molecular genetics 20.5 (2011): 905-916.
[9] Deng, San-Ming, et al. "The Notch ligand delta-like 3 promotes tumor growth and inhibits Notch signaling in lung cancer cells in mice." Biochemical and biophysical research communications 483.1 (2017): 488-494.
[10] Venkatesh, Deepak, et al. "RhoA-mediated signaling in Notch-induced senescence-like growth arrest and endothelial barrier dysfunction." Arteriosclerosis, thrombosis, and vascular biology 31.4 (2011): 876-882.
[11] Wang, Jianpeng, et al. "EGFL7 participates in regulating biological behavior of growth hormone–secreting pituitary adenomas via Notch2/DLL3 signaling pathway." Tumor Biology 39.7 (2017): 1010428317706203.
[12] Ding, Xiaojie, Fuyao Li, and Li Zhang. "Knockdown of Delta-like 3 restricts lipopolysaccharide-induced inflammation, migration and invasion of A2058 melanoma cells via blocking Twist1-mediated epithelial-mesenchymal transition." Life sciences 226 (2019): 149-155.
[13] Jia, Dongyu, et al. "NOTCH2/NOTCH3/DLL3/MAML1/ADAM17 signaling network is associated with ovarian cancer." Oncology letters 17.6 (2019): 4914-4920.
[14] Huang, Jianling, et al. "DLL3 is regulated by LIN28B and miR-518d-5p and regulates cell proliferation, migration and chemotherapy response in advanced small cell lung cancer." Biochemical and biophysical research communications 514.3 (2019): 853-860.
[15] Sabari, Joshua K., et al. "Unravelling the biology of SCLC: implications for therapy." Nature reviews Clinical oncology 14.9 (2017): 549-561.
[16] Vitorino, Philip, et al. "Rova-T enhances the anti-tumor activity of anti-PD1 in a murine model of small cell lung cancer with endogenous Dll3 expression." Translational oncology 14.1 (2021): 100883.
[17] Ayyanan, Ayyakannu, et al. "Increased Wnt signaling triggers oncogenic conversion of human breast epithelial cells by a Notch-dependent mechanism." Proceedings of the National Academy of Sciences 103.10 (2006): 3799-3804.
[18] Yan, S., et al. "Expression profile of Notch‐related genes in multidrug resistant K562/A02 cells compared with parental K562 cells." International journal of laboratory hematology 32.2 (2010): 150-158.
[19] Maemura, Kentaro, et al. "Delta-like 3 is silenced by methylation and induces apoptosis in human hepatocellular carcinoma." International journal of oncology 42.3 (2013): 817-822.
[20] Turchi, Laurent, et al. "Tumorigenic potential of miR‐18A* in glioma initiating cells requires NOTCH‐1 signaling." Stem Cells 31.7 (2013): 1252-1265.
[21] Jungk, Christine, et al. "Spatial transcriptome analysis reveals Notch pathway-associated prognostic markers in IDH1 wild-type glioblastoma involving the subventricular zone." BMC medicine 14.1 (2016): 1-16.
[22] Bauer, Todd M., et al. "ORAL02. 01: safety and efficacy of single-agent rovalpituzumab tesirine, a DLL3-targeted ADC, in recurrent or refractory SCLC: topic: medical oncology." Journal of Thoracic Oncology 11.11 (2016): S252-S253.
[23] Hamamoto, Hiroki, et al. "Delta-like 3 is silenced by HBx via histone acetylation in HBV-associated HCCs." Scientific reports 8.1 (2018): 1-11.
[24] Spino, Marissa, et al. "Cell surface Notch ligand DLL3 is a therapeutic target in isocitrate dehydrogenase–mutant glioma." Clinical Cancer Research 25.4 (2019): 1261-1271.
[25] Mizoguchi, Masahiro, et al. "Molecular characteristics of glioblastoma with 1p/19q co-deletion." Brain tumor pathology 29.3 (2012): 148-153.
[26] Song, Hai‑Yan, et al. "Expression of Notch receptors and their ligands in pancreatic ductal adenocarcinoma." Experimental and therapeutic medicine 16.1 (2018): 53-60.
[27] Kiniwa, Yukiko, et al. "Delta-like protein 3 promotes proliferation and migration of melanoma via MAPK activation." Journal of Dermatological Science 84.1 (2016): e176.
[28] Nakahara, Satoshi, et al. "AT-rich interaction domain-containing protein 3B has a potential to be a new stem cell marker of melanoma." Journal of Dermatological Science 84.1 (2016): e176-e177.
[29] Wang, Juan, et al. "Upregulated delta-like protein 3 expression is a diagnostic and prognostic marker in endometrial cancer: a retrospective study." Medicine 97.51 (2018).