- 移动端


武汉华美生物工程有限公司CUSABIO®品牌商
15 年
手机商铺
- NaN
- 0
- 0
- 2
- 2

推荐产品

公司新闻/正文
CDH1/E-cadherin:从黏附调控到肿瘤靶向治疗的核心靶点研究进展
356 人阅读发布时间:2026-03-20 11:52
E-cadherin由CDH1基因编码,是上皮细胞钙依赖性黏附分子,在维持上皮结构完整性、细胞间通讯与组织稳态中处于核心地位 [1,2]。CDH1功能受损常导致细胞间黏附松解并触发上皮-间充质转化(EMT)等关键表型变化,是多种肿瘤发生发展的重要驱动因素 [3]。本文围绕CDH1的结构与生理功能、多层次调控、关键通路、疾病谱和靶向药物最近研究进展进行结构化梳理,以期为您的研究带来帮助。
1. 背景与研究意义:CDH1为何是“黏附—信号—疾病”枢纽
2. CDH1/E-cadherin的结构与细胞-细胞黏附:从分子装配到力学耦联
3. 多层次调控机制:决定E-cadherin“表达水平与功能状态”的关键环节
4. 关键信号通路与细胞表型:从EMT到微环境适应
5. 相关疾病:从高外显率遗传综合征到多癌种与炎症/感染上皮病理
6. CDH1靶向药物最近研究进展
7. CDH1研究工具推荐:重组蛋白、抗体与ELISA试剂盒选型指南
1. 背景与研究意义:CDH1为何是“黏附—信号—疾病”枢纽
E-cadherin通过形成黏附连接(adherens junctions)将相邻细胞机械耦联,并与肌动蛋白骨架连接,参与细胞迁移、分化与信号转导 [2]。因此,E-cadherin通常被视为重要肿瘤抑制因子,其缺失可削弱细胞黏附并促进侵袭转移 [3]。除宏观组织结构外,E-cadherin还参与更精细的空间组织:在角质形成细胞中,E-cadherin缺失可造成微尺度细胞分离(micro-demixing),提示黏附在微观组织均质性/模式形成中具有“精细调参”作用 [4]。
临床上,CDH1致病性生殖系突变与遗传性弥漫性胃癌(HDGC)高度相关,携带者具有显著升高的弥漫性胃癌终生风险 [5-9],并且在特定人群(如新西兰毛利人)中被证实是早发弥漫性胃癌高发的重要遗传因素 [5]。除编码区突变外,下游调控序列缺失(如CDH1-TANGO6缺失)也可显著下调CDH1表达并导致极早发、高外显率的弥漫性胃癌 [10];在散发性胃癌中,CDH1遗传变异与表观遗传改变(如启动子甲基化)同样与致癌过程密切相关 [11]。此外,弥漫性胃癌中细胞-细胞/细胞-基质黏附依赖性的“逃逸”与RHO信号扰动(如RHOA突变或ARHGAP融合)相关,强调黏附改变与细胞内信号重塑存在协同关系 [12]。
值得注意的是,CDH1在不同肿瘤背景下可能呈现差异性作用。例如在乳腺癌中,CDH1可出现表达上调,并与分期、转移、干细胞特性及不良预后相关,提示其在特定背景下可能呈现促肿瘤效应 [13];而在治疗选择有限的三阴性乳腺癌中,CDH1缺陷较为常见,推动了“CDH1缺陷型肿瘤”靶向策略的探索 [14]。
2. CDH1/E-cadherin的结构与细胞-细胞黏附:从分子装配到力学耦联
E-cadherin是跨膜糖蛋白,胞外区由多个cadherin repeats构成,在钙离子存在下稳定并介导同型结合,是黏附连接形成的基础 [1]。其胞内区与β-catenin等衔接蛋白结合并连接肌动蛋白骨架,从而实现力学传递与组织尺度的机械耦联 [17,2]。相关生物物理模型将黏附复合物视作可传递阻力/张力的“弹簧样结构”,能够解释细胞极化、振荡动力学以及超细胞应力链等多细胞行为,提示“黏附—骨架”并非静态连接,而是双向耦合的动力系统 [2]。
结构层面的致病变异可直接破坏黏附功能。例如G212E错义突变会显著影响E-cadherin稳定性、定位与黏附能力,导致组织结构紊乱并削弱抗侵袭性 [19]。此外,E-cadherin功能也受复杂信号网络调控:如PAK5可参与维持细胞-细胞黏附完整性,提示黏附复合物并非“结构件”,而是受激酶网络持续调节的功能模块 [20]。
3. 多层次调控机制:决定E-cadherin“表达水平与功能状态”的关键环节
3.1 遗传与表观遗传调控
在HDGC及散发性胃癌中,CDH1可因编码区突变、调控序列缺失或启动子甲基化等机制导致表达下降或功能缺失 [5,10,11]。在炎症背景下,CDH1位点CpG甲基化增加也被报道与黏膜炎症相关联,提示慢性炎症可能通过表观遗传途径削弱上皮屏障 [24]。此外,乳腺癌中也存在CDH1异常甲基化与表达缺失的证据 [25]。
3.2 转录后调控与非编码RNA网络
多种miRNA/lncRNA被用于解释CDH1在不同肿瘤中的动态变化。例如,miR-92a-3p在胶质瘤及胶质瘤干样细胞中可通过靶向CDH1/β-catenin并影响Notch-1/Akt信号,参与肿瘤表型调控 [26];lncRNA SNHG1与hnRNPL形成复合体并共同调控CDH1,从而促进前列腺癌生长与转移 [32]。
3.3 翻译后修饰与蛋白互作
糖基化是影响E-cadherin活性与细胞行为的重要机制。在胰腺癌细胞中,ST3Gal III改变E-cadherin唾液酸化模式并降低细胞-细胞聚集能力,同时增强侵袭迁移相关信号(如FAK Tyr397磷酸化),提示“糖链-黏附-迁移”之间存在可观测的功能链条 [22]。互作层面,MCC蛋白可与E-cadherin及β-catenin互作并增强结直肠癌细胞黏附,提示黏附复合体稳定性还依赖肿瘤抑制网络的协同 [21]。
4. 关键信号通路与细胞表型:从EMT到微环境适应
CDH1缺陷最典型的后果是EMT相关表型增强。已有研究提出,SPHK1可通过促进自噬-溶酶体途径降解CDH1/E-cadherin从而诱导EMT,提示“代谢酶—自噬—黏附降解”的串联机制可能参与肝癌进展 [31]。另一方面,CDH1表达也与代谢重编程存在联系:E-cadherin可诱导丝氨酸合成以支持乳腺癌进展和转移,提示其在特定背景下可能通过代谢路径促进肿瘤适应 [28]。ZHX2缺陷可富集杂合型MET细胞并通过调控E-cadherin表达影响EMT/MET动态平衡,强调CDH1并非简单“开/关”,而可能参与多状态转换 [33]。
在通路层面,CDH1常与Wnt/β-catenin、PI3K/AKT/mTOR、MAPK/ERK及TGF-β/Smad等网络交织。既往研究在食管癌中讨论了Wnt/β-catenin与TGF-β-Smad通路的表观遗传失调及其对预后的影响 [3];在结直肠癌中,WNT通路组分也存在遗传与表观遗传改变并与微卫星不稳定性分层相关 [30]。
此外,CDH1异常还与上皮屏障破坏相关:在SARS-CoV-2感染的Caco-2肠上皮模型中,CDH1/E-cadherin表达与可溶性E-cadherin释放受到影响,并被讨论为肠道表现相关的潜在生理病理基础之一 [29]。
5. 相关疾病:从高外显率遗传综合征到多癌种与炎症/感染上皮病理
5.1 遗传性弥漫性胃癌(HDGC):以CDH1缺陷为核心的高风险疾病谱
HDGC是CDH1研究中最具“遗传—机制—临床管理”闭环特征的场景。现有研究强调:在新西兰毛利人群中,生殖系CDH1突变被证实显著贡献于早发弥漫性胃癌的高发生频率,提示其在特定遗传背景/人群结构下具有重要公共卫生意义 [5]。除经典编码区突变外,CDH1及其下游调控序列的联合缺失(CDH1-TANGO6缺失)可造成CDH1表达显著下降,并与极早发且高外显率弥漫性胃癌相关,提示“编码区之外的调控区域”同样可能决定疾病负担 [10]。
在散发性胃癌中,CDH1相关的遗传变异与表观遗传改变(例如调控区改变与启动子甲基化)被一并讨论为影响CDH1表达与致癌过程的重要因素,提示HDGC与散发性胃癌之间并非完全割裂,而可能在机制上存在“同轴不同强度”的连续谱 [11]。基于胃癌类器官模型的研究强调,疾病进展过程中可出现对细胞-细胞与细胞-基质黏附依赖性的“逃逸”,并伴随RHO信号扰动(如RHOA突变或ARHGAP融合)[12]。这类证据把“CDH1缺陷导致黏附失衡”与“细胞内信号重塑”连接起来,为理解弥漫性胃癌的侵袭性生物学行为提供机制线索 [12]。
5.2 乳腺癌:CDH1既可能是缺失驱动,也可能呈现情景依赖的“反直觉表达”
乳腺癌中CDH1呈现更强的异质性,CDH1生殖系突变与遗传性小叶型乳腺癌相关,被用于讨论遗传性肿瘤风险谱与遗传咨询路径 [9]。此外,家族性乳腺癌风险也被报道与CDH1等位基因SNP相关性有关 [27]。在部分研究中,CDH1可能出现表达上调,并与分期、转移、干细胞特性及不良预后相关,提示其在某些背景下可能呈现促肿瘤效应 [13]。这意味着在乳腺癌语境下,不能简单用“E-cadherin高=抑癌、低=促癌”概括,其临床解释往往需要结合肿瘤分型与分子网络 [13]。在三阴性乳腺癌等治疗选择有限场景中,CDH1缺陷更容易被纳入“可干预脆弱性”的讨论,并推动联合抑制策略探索 [14]。
5.3 结直肠癌:易感基因证据、黏附复合体稳态与炎症相关肿瘤免疫
结直肠癌相关证据链条更偏向“遗传易感—网络稳态—肿瘤免疫背景”,GWAS研究把CDH1纳入结直肠癌遗传易感基因图谱,提示其在群体层面具有风险相关性 [15]。MCC蛋白与E-cadherin/β-catenin互作可增强结直肠癌细胞黏附,强调CDH1相关表型不仅由单基因决定,也与黏附复合体伙伴蛋白网络有关 [21]。在炎症性肠病相关结直肠癌中,IBD相关基因被用于预后与肿瘤免疫含义分析,CDH1亦被纳入相关讨论框架 [16]。与此相呼应,结直肠癌中的WNT通路组分改变及其与微卫星不稳定性分层的关系,为解释上皮稳定性破坏与信号重编程提供了宏观通路层面的背景 [30]。
5.4 食管癌与癌前病变:CDH1/CTNNB1表达下降与转移、预后相关
在食管癌中,CDH1或CTNNB1表达降低与淋巴结转移及不良预后相关,这一观察把“黏附复合体失衡”与更不利的临床结局联系起来 [17]。同时,关于Wnt/β-catenin与TGF-β-Smad通路的表观遗传失调被用于解释食管癌的预后差异,提示CDH1改变往往与更大范围的通路层级异常并行出现 [3]。
在癌前病变层面,口腔扁平苔藓中EMT相关蛋白(含E-cadherin)的表达改变提示细胞连接紊乱可能与病变演进相关,为“黏附改变的早期提示意义”提供了证据线索 [18]。
5.5 炎症与感染相关上皮屏障:甲基化改变与可溶性E-cadherin释放
CDH1不仅与肿瘤相关,也与上皮屏障状态密切相关:在炎症背景下,CDH1位点CpG甲基化增加与黏膜炎症相关联,提示慢性炎症可能通过表观遗传途径影响上皮黏附与屏障稳态 [24]。在SARS-CoV-2感染的Caco-2肠上皮模型中,CDH1/E-cadherin表达及可溶性E-cadherin释放发生变化,被用于讨论肠道表现相关的病理基础之一 [29]。与检测相关的证据还包括:尿液中可溶性E-cadherin片段升高被认为可能反映上皮肿瘤细胞的剪切/脱落过程,为非侵入性标志物提供线索 [23]。
6. CDH1靶向药物最近研究进展
目前靶向CDH1的药物研发主要处于临床前及早期发现阶段,涵盖小分子化药、生物药、外泌体等多种类型。主要探索方向包括脑恶性胶质瘤、乳腺癌及多囊疾病等,涉及沈阳药科大学、哈佛大学、四川省肿瘤医院等多家机构。
| 药物 | 类型 | 适应症 | 研发阶段 | 研发机构 |
|---|---|---|---|---|
| AL-GDa62 | 小分子抑制剂 | CDH1缺陷型胃癌 | 临床前 | 新西兰奥塔哥大学 |
| Dasatinib | 多激酶抑制剂 | CDH1缺陷型肿瘤 | 临床前 | 多个研究机构 |
| FAK抑制剂 + ROS1抑制剂 | 联合疗法 | CDH1缺陷型癌症 | 临床前 | 多个研究机构 |
| 外泌体递送CDH1 mRNA | 基因治疗 | 上皮屏障损伤修复 | 探索阶段 | 国内科研机构 |
7. CDH1研究工具推荐:重组蛋白、抗体与ELISA试剂盒选型指南
华美生物提供CDH1重组蛋白、抗体及ELISA试剂盒产品,助力您进行相关机制研究及靶向药物开发。
● CDH1 重组蛋白
Recombinant Human Cadherin-1(CDH1),partial (Active); CSB-MP005034HU1
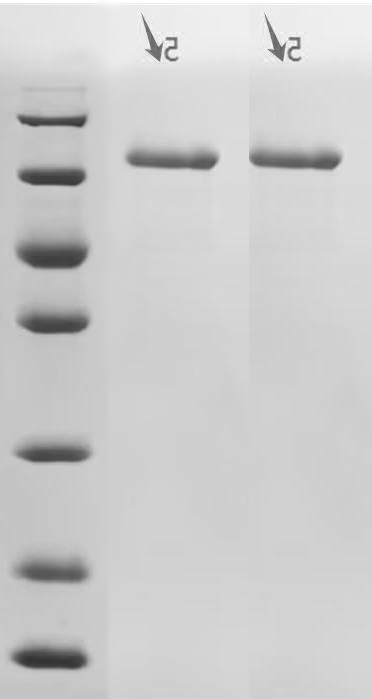
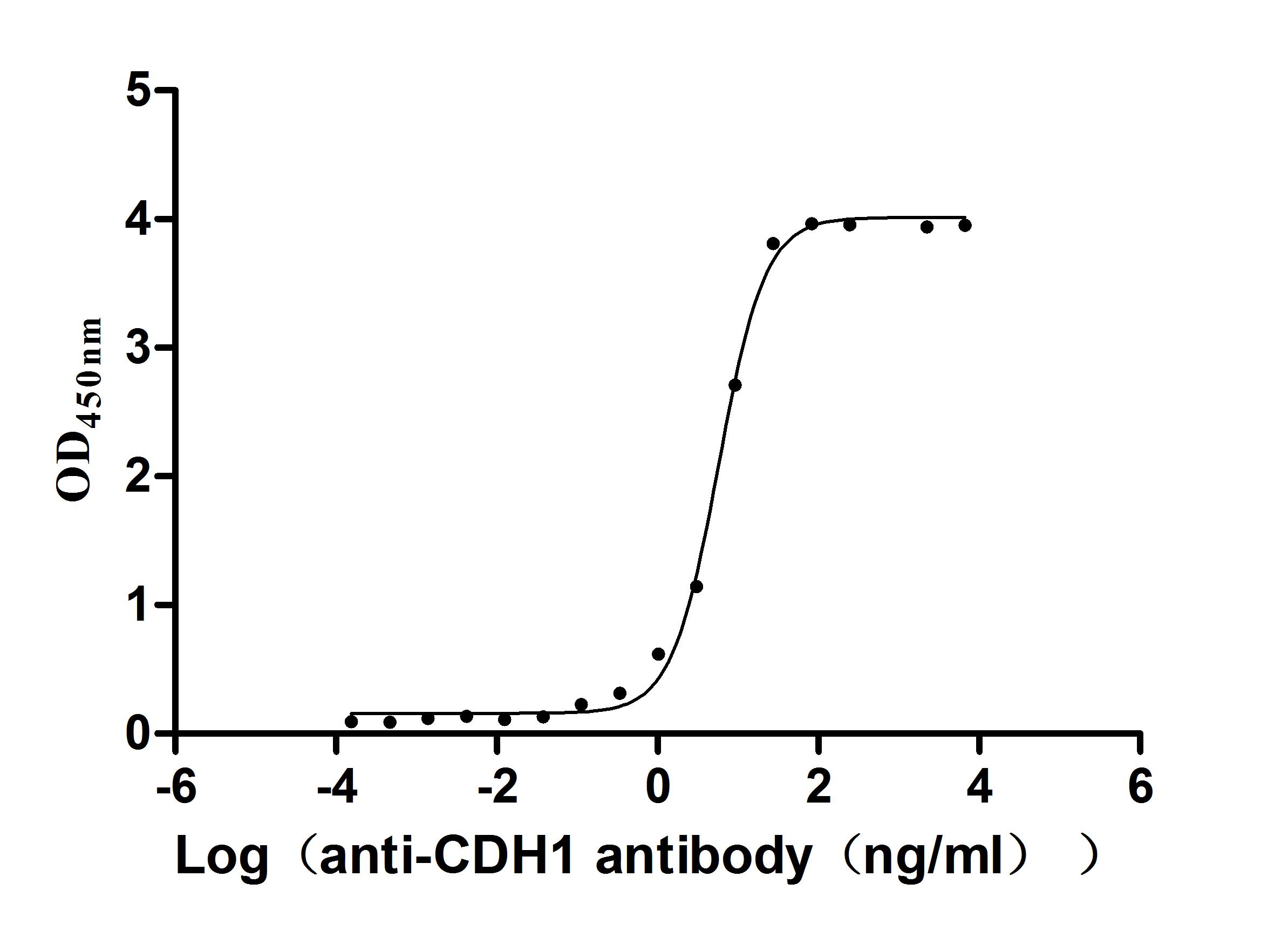
Recombinant Macaca fascicularis Cadherin-1 (CDH1), partial (Active); CSB-MP5601MOV
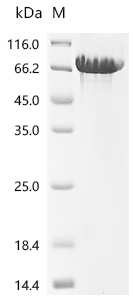
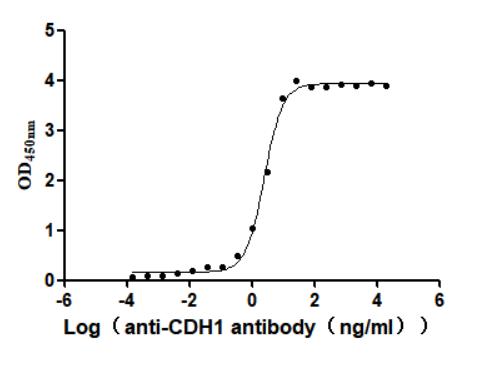
● CDH1 抗体
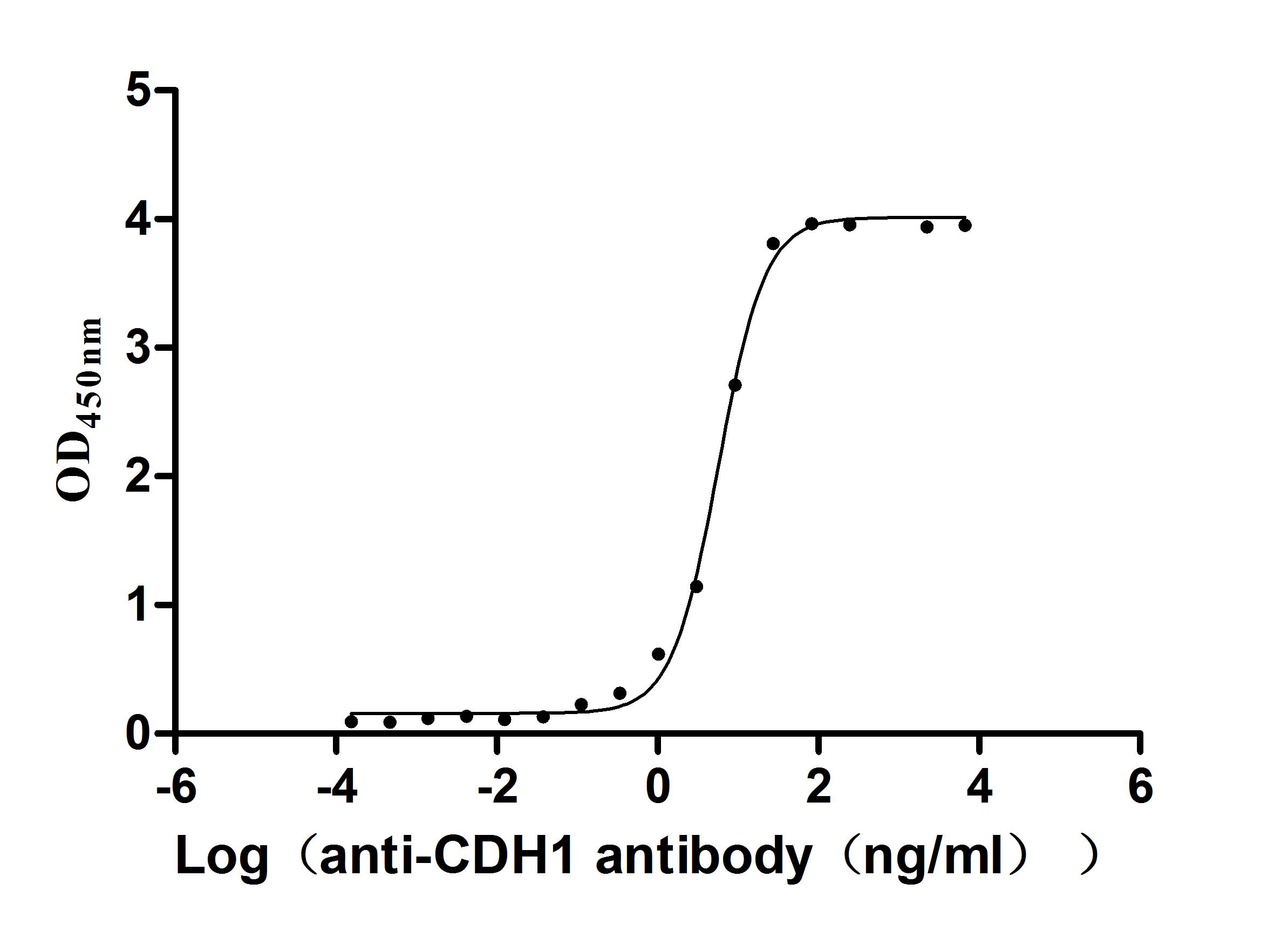
CDH1 Recombinant Monoclonal Antibody; CSB-RA005034MA1HU
CDH1 Recombinant Monoclonal Antibody
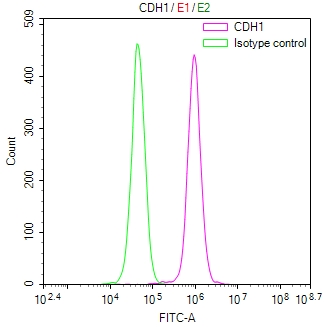
CSB-RA576116A0HU
CDH1 Recombinant Monoclonal Antibody
CSB-RA005034MA2HU
CDH1/CDH2/CDH3/CDH5/CDH4 Recombinant Monoclonal Antibody
CSB-RA291625A0HU
● CDH1 ELISA 试剂盒
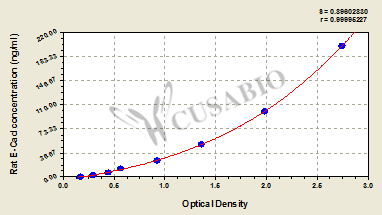
Rat Epithelial-Cadherin,E-Cad ELISA Kit
CSB-E07308r
Human E-Cadherin,E-Cad ELISA Kit
CSB-E04519h
[1] Anna Zhigun, Mabel Lizzy Rajendran.(2023). Modelling non-local cell-cell adhesion: a multiscale approach.
[2] XinXin Du, Ido Lavi, Michael J. Shelley.(2025). Collective multicellular patterns arising from cadherin-linked cytoskeletal domains.
[3] Virendra Singh, A. Singh, I. Sharma, L. Singh, J. Sharma, B. B. Borthakar, A. Rai, A. Kataki, S. Kapur, S. Saxena.(2019). Epigenetic deregulations of Wnt/β-catenin and transforming growth factor beta-Smad pathways in esophageal cancer: Outcome of DNA methylation.
[4] Preeti Sahu, Daniel M. Sussman, Matthias Rubsam, Aaron F. Mertz, Valerie Horsley, Eric R. Dufresne, Carien M. Niessen, M. Cristina Marchetti, M. Lisa Manning, J. M. Schwarz.(2019). Small-scale demixing in confluent biological tissues.
[5] C. Hakkaart, L. Ellison-Loschmann, R. Day, A. Sporle, Jonathan Koea, Pauline Harawira, Soo Cheng, M. Gray, Tracey Whaanga, N. Pearce, P. Guilford.(2018). Germline CDH1 mutations are a significant contributor to the high frequency of early-onset diffuse gastric cancer cases in New Zealand Māori.
[6] I. Chen, Lesley Mathews-Greiner, Dandan Li, A. Abisoye-Ogunniyan, Satyajit Ray, Yansong Bian, V. Shukla, Xiaohu Zhang, Rajarashi Guha, Craig J. Thomas, B. Gryder, A. Zacharia, J. Beane, S. Ravichandran, M. Ferrer, U. Rudloff.(2017). Transcriptomic profiling and quantitative high-throughput (qHTS) drug screening of CDH1 deficient hereditary diffuse gastric cancer (HDGC) cells identify treatment leads for familial gastric cancer.
[7] Nicola Bougen-Zhukov, Lyvianne Decourtye-Espiard, Wilson Mitchell, Kieran Redpath, J. Perkinson, T. Godwin, M. Black, P. Guilford.(2022). E-Cadherin-Deficient Cells Are Sensitive to the Multikinase Inhibitor Dasatinib.
[8] T. Godwin, S. Kelly, Tom Brew, Nicola Bougen-Zhukov, A. Single, Augustine Chen, Cassie E Stylianou, L. Harris, Sophie K Currie, Bryony J. Telford, Henry Beetham, G. Evans, M. Black, P. Guilford.(2018). E-cadherin-deficient cells have synthetic lethal vulnerabilities in plasma membrane organisation, dynamics and function.
[9] G. Corso, M. Intra, C. Trentin, P. Veronesi, V. Galimberti.(2016). CDH1 germline mutations and hereditary lobular breast cancer.
[10] C. São José, J. Garcia-Pelaez, Marta Ferreira, O. Arrieta, Ana André, Nelson Martins, Samantha Solís, B. Martínez-Benítez, M. Ordoñez-Sánchez, M. Rodríguez-Torres, Anna K. Sommer, Iris B A W Te Paske, C. Caldas, M. Tischkowitz, María Teresa Tusié, Stefan Gabriel Sérgio Richarda M. Gareth Susana José Luzi Aretz Capella Castedo de Voer Evans Fernandes Garc, S. Aretz, G. Capellá, S. Castedo, Richarda M. de Voer, Gareth Evans, Susana Fernandes, J. Garcia-Pelaez, L. Garrido, E. Holinski-Feder, N. Hoogerbrugge, D. Huntsman, A. Jahn, C. Kets, A. Laner, M. Ligtenberg, Andrea Meinhardt, A. Mensenkamp, Carla Oliveira, S. Peters, I. Quintana, E. Schröck, Anna K. Sommer, I. Spier, L. Spruijt, V. Steinke-Lange, I. T. Paske, M. Tischkowitz, L. Valle, R. S. van der Post, Y. V. van Herwaarden, W. V. van Zelst-Stams, D. William, N. Hoogerbrugge, German Demidov, Richarda M. de Voer, S. Laurie, Carla Oliveira.(2023). Combined loss of CDH1 and downstream regulatory sequences drive early-onset diffuse gastric cancer and increase penetrance of hereditary diffuse gastric cancer.
[11] G. Tedaldi, C. Molinari, C. São José, Rita Barbosa-Matos, A. André, R. Danesi, V. Arcangeli, M. Ravegnani, L. Saragoni, P. Morgagni, F. Rebuzzi, M. Canale, S. Pignatta, Elisa Ferracci, G. Martinelli, G. Ranzani, Carla Oliveira, D. Calistri, P. Ulivi.(2021). Genetic and Epigenetic Alterations of CDH1 Regulatory Regions in Hereditary and Sporadic Gastric Cancer.
[12] Yin Tong, P. Cheng, Chung Sze Or, Sarah S K Yue, H. Siu, S. Ho, S. Law, W. Tsui, D. Chan, Stephanie Ma, Siu Po Lee, Annie S. Y. Chan, April S Chan, Shui Wa Yun, Ho Sang Hui, S. Yuen, S. Leung, Helen H. N. Yan.(2022). Escape from cell-cell and cell-matrix adhesion dependence underscores disease progression in gastric cancer organoid models.
[13] D. Xie, Yiyu Chen, Xue Wan, Jingyuan Li, Qin Pei, Yanan Luo, Jinbo Liu, Ting Ye.(2022). The Potential Role of CDH1 as an Oncogene Combined With Related miRNAs and Their Diagnostic Value in Breast Cancer.
[14] Jiaming Gao, Yunying Yao, Zaiqi Wang, Baoyuan Zhang, R. Ren.(2023). Abstract 5821: Synergism of FAK and ROS1 inhibitors in the treatment of CDH1-deficient cancers mediated by FAK-YAP-TRX signaling.
[15] Da-Tian Bau, Ting-Yuan Liu, Jai-Sing Yang, William Tzu-Liang Chen, Chia‐Wen Tsai, Wen‐Shin Chang, Tao-Wei Ke, Chi-Chou Liao, Yu-Chia Chen, Yen-Ting Chang, Fuu-Jen Tsai.(2024). Characterizing Genetic Susceptibility to Colorectal Cancer in Taiwan Through Genome‐Wide Association Study.
[16] Di Wang, Biao Xie.(2022). Prognostic and tumor immunity implication of inflammatory bowel disease-associated genes in colorectal cancer.
[17] H. Ishiguro, T. Wakasugi, Yukio Terashita, N. Sakamoto, Tatsuya Tanaka, K. Mizoguchi, H. Sagawa, T. Okubo, H. Takeyama.(2016). Decreased expression of CDH1 or CTNNB1 affects poor prognosis of patients with esophageal cancer.
[18] L. Hämäläinen, Y. Soini, S. Pasonen-Seppänen, M. Siponen.(2019). Alterations in the expression of EMT-related proteins claudin-1, -4 and -7, E-cadherin, TWIST1 and ZEB1 in oral lichen planus..
[19] J. Figueiredo, F. Mercadillo, Soraia Melo, A. Barroso, Margarida Gonçalves, J. Diaz-Tasende, Patrícia Carneiro, L. Robles, F. Colina, C. Ibarrola, J. Perea, Eurico Morais-de-Sá, R. Seruca, M. Urioste.(2021). Germline CDH1 G212E Missense Variant: Combining Clinical, In Vitro and In Vivo Strategies to Unravel Disease Burden.
[20] A. F. Ismail, Sevil Oskay Halacli, N. Babteen, M. De Piano, T. Martin, W. Jiang, Muhammad S. Khan, P. Dasgupta, C. Wells.(2017). PAK5 mediates cell: cell adhesion integrity via interaction with E-cadherin in bladder cancer cells..
[21] F. Benthani, F. Benthani, D. Herrmann, D. Herrmann, P. N. Tran, P. N. Tran, L. Pangon, L. Pangon, M. Lucas, A. Allam, N. Currey, S. Al‐Sohaily, Marc Giry-Laterrière, J. Warusavitarne, P. Timpson, P. Timpson, M. R. Kohonen-Corish, M. R. Kohonen-Corish, M. R. Kohonen-Corish.(2018). ‘MCC’ protein interacts with E-cadherin and β-catenin strengthening cell–cell adhesion of HCT116 colon cancer cells.
[22] Sònia Bassagañas, S. Carvalho, A. M. Dias, Marta Pérez-Garay, M. R. Ortiz, J. Figueras, C. Reis, S. Pinho, R. Peracaula.(2014). Pancreatic Cancer Cell Glycosylation Regulates Cell Adhesion and Invasion through the Modulation of α2β1 Integrin and E-Cadherin Function.
[23] M. Katayama, S. Hirai, M. Yasumoto, K. Nishikawa, S. Nagata, M. Otsuka, K. Kamihagi, I. Kato.(1994). Soluble fragments of e-cadherin cell-adhesion molecule increase in urinary-excretion of cancer-patients, potentially indicating its shedding from epithelial tumor-cells..
[24] Charles de Ponthaud, Solafah Abdalla, M. Belot, Xiaojian Shao, Christophe Penna, A. Brouquet, Pierre Bougnères.(2024). Increased CpG methylation at the CDH1 locus in inflamed ileal mucosa of patients with Crohn disease.
[25] Asia Asiaf, S. T. Ahmad, S. Aziz, A. Malik, Zubaida Rasool, A. Masood, M. Zargar.(2014). Loss of expression and aberrant methylation of the CDH1 (E-cadherin) gene in breast cancer patients from Kashmir..
[26] Hang Song, Yao-quan Zhang, Na Liu, Sheng Zhao, Y. Kong, Liudi Yuan.(2016). miR-92a-3p Exerts Various Effects in Glioma and Glioma Stem-Like Cells Specifically Targeting CDH1/β-Catenin and Notch-1/Akt Signaling Pathways.
[27] M. Bagherpour, Kamelia Gharibzad, H. Rassi.(2018). Association of CDH1 and TERT Single-Nucleotide Polymorphisms with Susceptibility to Familial Breast Cancer Risk..
[28] Geonhui Lee, Claudia Wong, Anna Cho, Junior J West, Ashleigh J. Crawford, Gabriella C. Russo, B. R. Si, Jungwoo Kim, Lauren Hoffner, Cholsoon Jang, Moonjung Jung, Robert D Leone, K. Konstantopoulos, Andrew J Ewald, Denis Wirtz, Sangmoo Jeong.(2024). E-cadherin Induces Serine Synthesis to Support Progression and Metastasis of Breast Cancer..
[29] I. O. Osman, Clémence Garrec, Gabriel Augusto Pires de Souza, A. Zarubica, Djamal Brahim Belhaouari, J. Baudoin, H. Lepidi, J. Mege, B. Malissen, B. Scola, C. Devaux.(2022). Control of CDH1/E-Cadherin Gene Expression and Release of a Soluble Form of E-Cadherin in SARS-CoV-2 Infected Caco-2 Intestinal Cells: Physiopathological Consequences for the Intestinal Forms of COVID-19.
[30] L. Thorstensen, G. E. Lind, T. Løvig, C. B. Diep, G. Meling, T. Rognum, R. Lothe.(2005). Genetic and epigenetic changes of components affecting the WNT pathway in colorectal carcinomas stratified by microsatellite instability..
[31] Hong Liu, Yan Ma, Hong-wei He, Wu-li Zhao, R. Shao.(2017). SPHK1 (sphingosine kinase 1) induces epithelial-mesenchymal transition by promoting the autophagy-linked lysosomal degradation of CDH1/E-cadherin in hepatoma cells.
[32] Xiao Tan, Wenbin Chen, D. Lv, Tao-wei Yang, Kai-hui Wu, Li-bin Zou, Junqi Luo, Xu-min Zhou, Guo-chang Liu, F. Shu, X. Mao.(2020). LncRNA SNHG1 and RNA binding protein hnRNPL form a complex and coregulate CDH1 to boost the growth and metastasis of prostate cancer.
[33] Yan He, Qimin Zhang, Yuan-Xiao Chen, Yingjian Wu, Yuan Quan, Weihua Chen, Jing Yao, Peijing Zhang.(2023). ZHX2 deficiency enriches hybrid MET cells through regulating E-cadherin expression.